
Cialis ist bekannt für seine lange Wirkdauer von bis zu 36 Stunden. Dadurch unterscheidet es sich deutlich von Viagra. Viele Schweizer vergleichen daher Preise und schauen nach Angeboten unter dem Begriff cialis generika schweiz, da Generika erschwinglicher sind.
Doi:10.1016/j.phytochem.2004.02.009

Phytochemistry 65 (2004) 865–873
Variation in pyrrolizidine alkaloid patterns of Senecio jacobaea
Mirka Macel, Klaas Vrieling, Peter G.L. Klinkhamer
Leiden University, Institute for Biology, Plant Ecology, PO Box 95162300 RA Leiden, The Netherlands
Received 30 July 2003; accepted 10 February 2004
We studied the variation in pyrrolizidine alkaloid (PA) patterns of lab-grown vegetative plants of 11 European Senecio jacobaea
populations. Plants were classified as jacobine, erucifoline, mixed or senecionine chemotypes based on presence and absence of thePAs jacobine or erucifoline. Due to the presence of jacobine, total PA concentration in jacobine chemotypes was higher than inerucifoline chemotypes. Both relative and absolute concentrations of individual PAs differed between half-sib and clonal families,which showed that variation in PA patterns had a genetic basis. Within most populations relative abundance of PAs varied con-siderably between individual plants. Most populations consisted either of the jacobine chemotype or of the erucifoline chemotype,sometimes in combination with mixed or senecionine chemotypes.
# 2004 Elsevier Ltd. All rights reserved.
Keywords: Senecio jacobaea; Genetic variation; Geographic distribution; Pyrrolizidine alkaloids; Chemotypes; Jacobine; Erucifoline
the jacobine type and the erucifoline type, based on PAcontent in inflorescences of plants collected in the field.
Plants from the genus Senecio (Asteraceae) are known
Jacobine types were characterized by the PAs jacobine
for the production of a wide variety of pyrrolizidine
and jacozine and lacked erucifoline, while the erucifo-
alkaloids (PAs) ) that have
line types contained erucifoline and acetylerucifoline but
hepatotoxic and carcinogenic properties
hardly any jacobine (see for structures). The two
In Senecio species PAs are pro-
chemotypes did not differ in concentrations of other
duced in the roots as senecionine N-oxides (
PAs such as senecivernine, senecionine, integerrimine
and seneciphylline. Only rarely an intermediate chemo-
N-oxide is transported via the phloem to the above
type was found that contained both jacobine and eruci-
ground plant organs (where it is
foline. The patterns described by are
transformed into several related PAs (Hartmann and
interesting in relation to the evolution of diversity of
Dierich, 1998). The transformation of senecionine into
alkaloids. However, generally only one plant per popu-
related PAs differs between Senecio species and hence
lation was sampled and collected in its natural habitat.
species-specific PA patterns are produced (Hartmann
Therefore, it is not possible to disentangle environmen-
and Dierich, 1998). Senecio jacobaea can contain more
tally induced patterns and genetic effects. Here, we
than 10senecionine related alkaloids
investigated the PA patterns in leaves of vegetative
PA concentration in S. jacobaea is partly geneti-
plants from 10European populations of S. jacobaea. To
cally determined (
determine whether the different PA patterns have a
described two PA chemotypes for S. jacobaea,
genetic basis, we studied these patterns in cloned S.
jacobaea genotypes and half-sib families grown underidentical controlled conditions to minimize a possible
* Present address: University of Fribourg, Department of Biology,
effect of environment. To determine the variation within
Ecology and Evolution, Chemin du Muse´e 10, CH-1700 Fribourg,
and between the different populations, PAs from 5
Switzerland. Tel.: +41-26-3008849; fax: +41-26-3009698.
E-mail address: (M. Macel).
plants of every population were analyzed.
0031-9422/$ - see front matter # 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.phytochem.2004.02.009
M. Macel et al. / Phytochemistry 65 (2004) 865–873
Fig. 1. Structures and codes of PAs found in S. jacobaea. sv=senecivernine; sn=senecionine; sp=seneciphylline; in=integerrimine; jb=jacobine;jz=jacozine; jl=jacoline; jn=jaconine; er=erucifoline; fl=eruciflorine.
2. Results and discussion
erucifoline (KW, df=9, for all PAs: P < 0.005). Varia-tion in PA composition within clonal families was small
2.1. Variation in PA composition within clonal families
compared to variation among these families indicating astrong genetic component. Inheritance of PA patterns
To show that variation in PA composition among
could further be investigated by crossing genotypes with
plants is genetically determined, we studied the varia-
a different PA composition.
tion in PA profiles within 10clonal families of S. jaco-baea (16–34 plants per family). Each clonal family was
2.2. Variation in PA composition between half-sib
from a different population (). In some plants 6
senecionine related PAs were found while other plantsonly contained two PAs. The PA composition (relative
We analyzed the PA composition of offspring from 25
abundance) of each clonal family is shown in
plants in two areas (in total 50half-sib families) of the
Clonal families differed in relative percentages of sene-
population in Meijendel (The Netherlands). All 413
cionine, seneciphylline, integerrimine, jacobine and eru-
plants analyzed, except for one plant, contained mainly
cifoline (KW, df=9, for all PAs: P < 0.005). The clonal
jacobine, the percentage of jacobine ranged from 41 to
families also differed in absolute concentrations of
100% of total PA. One plant contained mainly jacoline
senecionine, seneciphylline, integerrimine, jacobine and
(45%). The percentage of erucifoline ranged from 0to
Table 1Origin of S. jacobaea plants used in this study. Source indicates number of plants of which seeds were used in our study
Amsterdam, The Netherlands
Burghaamstede, The Netherlands
Driel, The Netherlands
Slenaken, The Netherlands
Meijendel, The Netherlands
Mavellier, Switzerland
Sundsvall, Sweden
Ejby Adal, Denmark
Schiermonnikoog, The Netherlands

M. Macel et al. / Phytochemistry 65 (2004) 865–873
Fig. 2. PA composition of S. jacobaea clonal families in average relative abundance of PAs ( � S.D.). Most clonal families that contained jacobinealso contained small amounts of erucifoline. Origin is indicated in the right corner. Average PA concentration per clonal family ( � SE) is given.
n=16–34.

M. Macel et al. / Phytochemistry 65 (2004) 865–873
plants from the same population that we analyzed weresometimes half-sibs (and therefore the actualvariation in PA composition in a population might beeven greater than is shown by our data.
All plants contained the PAs senecionine or seneci-
phylline. In contrast to the chemotypes described by, plants that contained jacobine had
Fig. 3. PA composition of S. jacobaea from Meijendel in relativeabundance of PAs ( � S.D.). n=413.
often also small amounts of erucifoline (Thejacobine types described by com-
19% of total PA. shows the mean relative abun-
pletely lacked erucifoline. Here, we will consider (1)
dance (%) of individual PAs over all 413 plants. The
plants with mainly jacobine and no or little erucifoline
percentage of senecionine, seneciphylline, integerrimine,
as jacobine chemotypes, (2) plants that contained both
jacobine and jacozine differed among the families (KW,
jacobine and erucifoline in similar amounts as mixed
df=49, for all PAs P < 0.005). Percentages erucifoline
chemotypes and (3) plants without jacobine but with
and jacoline did not differ among the families (er: KW
erucifoline as erucifoline chemotypes. In addition, some
w2=61.51, df=49, ns ; jl: KW w2=45.93, df=49, ns). In
plants had only trace amounts of erucifoline and con-
absolute concentrations senecionine, seneciphylline,
tained no jacobine. We classified these plants as sene-
integerrimine, jacobine and jacozine also differed
cionine chemotypes. shows the PA composition
between the families (KW, df=49, for all PAs
of these chemotypes.
P < 0.002). Jacoline and erucifoline concentrations did
Most populations consisted of either erucifoline che-
not differ between the families (KW, ns). Families also
motypes or jacobine chemotypes, sometimes in combi-
did differ in their total PA concentration ranging from
nation with mixed or senecionine chemotypes (,
1.57 mg/g (se � 0.30) to 5.51 mg/g (se � 0.97) (ANOVA
). In five of the ten populations we sampled (DRL,
F=2.64, df=49, P < 0.001). PA concentration did not
SUN, FIL, CHE, BUR) mixed chemotypes were found
differ between plants from the two areas in Meijendel
(Only in one population, CHE, plants of
(ANOVA, F=1.79, df=1, P=0.18). These results show
pure jacobine, mixed and pure erucifoline chemotype
that although all plants contained mainly jacobine,
were found It is possible, however, that more
considerable (genetic) variation in both PA composition
‘mixed' populations exists because we may not have a
and concentration existed within the Meijendel popula-tion.
Table 2PA composition of plants from three S. jacobaea populations as an
2.3. Variation within and between populations
example for the variation within and between populations
From 10European S. jacobaea populations
Alkaloid (% relative abundance)
4–5 plants were analyzed for their PA composition. Ten
senecionine related PAs were found in some plants whileother plants contained only two or three of these PAs.
shows the average PA composition of the popu-
lations. Populations differed significantly in the relative
abundance of seneciphylline (KW w2=24.53, df=9,
P < 0.001). Relative abundance of all other PAs did not
differ between populations (KW, df=9, all P > 0.05). In
absolute concentrations only jacobine differed among
the populations (KW w2=24.98, df=9, P < 0.005), con-
centrations of the other PAs did not differ between the
populations (KW, df=9, all P > 0.05). Although a rela-
tively small number of plants was used of each popula-
tion, variation in PA composition within populationswas considerable and this variation confirms the more
The AMS population consists of jacobine and senecionine chemo-types, SLK population consists of erucifoline chemotypes only and the
extensive data from the Meijendel population above. As
CHE popopulation consists of jacobine, erucifoline and mixed che-
an example, shows the variation in PA compo-
motypes based on the relative abundance of individual PAs. tr=trace
sition of individual plants in three populations. The
amounts < 1% of total PA

M. Macel et al. / Phytochemistry 65 (2004) 865–873
Fig. 4. PA compositions of S. jacobaea from European populations given as average relative abundance of PAs ( � S.D.). Population is indicated inthe right corner. Average PA concentration per populations in mg/g dry weight ( � SE) is given. Letters indicate chemotypes of individual plants:S=senecionine type, E=erucifoline type, J=jacobine type, M=mixed type n=4–5.

M. Macel et al. / Phytochemistry 65 (2004) 865–873
Fig. 5. PA composition of the chemotypes in relative abundance of PAs ( � SE).
complete overview of all the chemotypes present in a
types (). Among the plants from the populations,
population due to the limited number of genotypes of
the mixed chemotype and senecionine chemotype had
most populations. suggested that the
intermediate PA concentrations (In the clonal
distribution of the erucifoline chemotype was limited to
families, senecionine chemotypes also had a lower total
the Eastern and Southern Europe. However, in our
PA concentration compared to the jacobine chemotypes
study, plants of Northern European populations (Swe-
(). The higher total PA concentration of the jaco-
den, Denmark) were erucifoline chemotypes (
bine chemotype is due to the ‘‘addition'' of jacobine
Furthermore, plants of some populations from The
(If we compared the total concentration minus
Netherlands also were erucifoline chemotypes. The PA
jacobine we found no significant difference between the
patterns found under uniform conditions may perhaps
chemotypes for the populations (ANOVA, F=1.45,
not accurately reflect differences in PA patterns between
df=4, P=0.281). For the clones the total PA concen-
populations in the field because plants may show differ-
tration without jacobine was significantly higher in the
ent norms of reaction for e.g. nutrient concentration,
erucifoline and senecionine chemotypes (KW w2=23.93,
water availability, day length. Our data do show that
df=2, P < 0.001).
not only South-Eastern populations are geneticallyequipped to produce erucifoline chemotypes.
2.5. Natural selection?
The total PA concentration of the jacobine chemo-
types in both the clones and the populations was higher
Our data for the clonal families show that differences
than the PA concentration of the erucifoline chemo-
in PA composition between populations of S. jacobaea
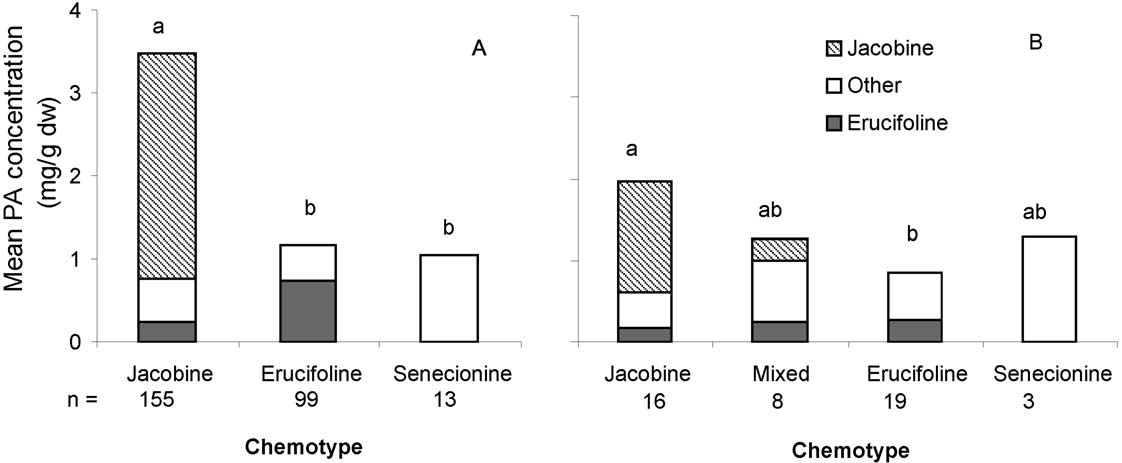

M. Macel et al. / Phytochemistry 65 (2004) 865–873
have a genetic basis. The variation in PA composition
chemical range as studied in this paper, showed that
between individual plants within the populations was
there was considerable variation among different popu-
considerable. In the population of Meijendel, half-sib
lations. This molecular variation was not linked to the
families differed in PA composition and concentration.
distribution of alkaloid patterns, although one marker
Molecular genetic analyses (AFLP) of European S.
was directly linked with presence of jacobine (K. van
jacobaea populations, with similar geographic and bio-
den Hof, J. Joshi and K. Vrieling, unpublished data).
Fig. 6. Geographic distribution for the chemotypes of generative plants and vegetative plants of S. jacobaea in Europe.
Fig. 7. Average total PA concentrations of the chemotypes from plants of (A) clones or (B) populations. Letters indicate significant differences intotal PA concentration, Populations: ANOVA, Bonferroni post-hoc tests, Clonal families: Kruskal–Wallis, post-hoc Mann–Whitney tests withBonferroni correction.
M. Macel et al. / Phytochemistry 65 (2004) 865–873
This seems to indicate that the geographical distribution
DB-17 80kPa, split mode 1-20
, detectors: FID and
of the chemotypes in Europe is not caused by common
NPD. Structure of the PAs was verified by GC–MS
ancestry but by similar selection pressures in a certain
(Total PA concentration was deter-
region. The genetic variation in PA composition pro-
mined spectrophotometrically, by a method modified
vides a basis upon which natural selection, by e.g. her-
after Heliotrine (Latoxan) was used as
bivores or pathogens, may act. It is unlikely that the
ref. compound. The concentration of individual PAs
specialist herbivore Tyria jacobaeae plays a role in the
was calculated by total PA concentration�fraction of
evolution of the different chemotypes because larval
individual PA measured by GC analysis.
performance and oviposition preference of this specia-list moth was not affected by chemotype of S. jacobaea
3.4. Statistical analysis
However, other herbivores may
Statistics were performed in SPSS 8.0(SPSS Inc,
have been sensitive to difference in PA composition. It
1998). Total PA concentrations of the 413 plants from
has been shown that the relative effect of a PA can differ
the half-sib families in Meijendel were log-transformed,
between insect species (and therefore dif-
after log-transformation differences in PA concentration
ferences in herbivore community in S. jacobaea popula-
among families were tested with a one-way ANOVA.
tions could have led to the evolution of the different
Differences between individual PAs were tested with a
Kruskal–Wallis test. Differences in PA composition andconcentrations among the clonal families and popula-tions were also tested with a Kruskal–Wallis test.
3.1. Growth conditions
All plants were grown in 50/50 dune sand/peat mix-
We are most grateful to Dr. Jens Hagen, Dr. L. Witte
ture in 11 cm diameter pots in a growth chamber: pho-
and Prof. Dr. T. Hartmann for their help with the GC
toperiod 8 h light: 16 h dark, 20 �C day/15 �C night,
and GC–MS analysis. We thank Helene de Vos and
relative humidity 70%. After two months in the pots the
Karin van Veen- van Wijk for their technical assistance
plants were given ample nutrients.
and Martin Brittijn for the artwork. Urs Schaffner,Stefan Andersson, Els Schlatmann, Henk van Dijk,
3.2. Clonal families, half-sib families and populations
Nico de Boer and the Botanical Garden in Copenhagengenerously provided us with seeds from the different
Clonal families: We used 16–34 plants per clonal
populations. We thank Nicole van Dam, Ed van der
family. A clonal family consisted of one randomly
Meijden and two anonymous referees for their valuable
selected genotype of a population. This genotype was
comments on earlier drafts of the manuscript.
propagated via tissue culture. Plants were grown for twomonths in pots before analyzing the PA patterns.
Half-sib families: We collected seeds from S. jacobaea
plants in two areas of the Meijendel population. In each
Cheeke, P.R., 1988. Toxicity and metabolism of pyrrolizidine alka-
area we sampled 25 plants. We analyzed 4–10seedlings
loids. J. Anim. Sci. 66, 2343–2350.
per plant for their PA profile, in total 413 plants. Plants
Hartmann, T., Dierich, B., 1998. Chemical diversity and variation of
were grown from seed for two months before analysis.
pyrrolizidine alkaloids of the senecionine type: biological need or
Populations: Plants were grown from seed for 5
coincidence? Planta 206, 443–451.
Hartmann, T., Ehmke, A., Eilert, U., Von Borstel, K., Theuring, C.,
months and then harvested for PA analysis.
1989. Sites of synthesis, translocation and accumulation of pyrroli-zidine alkaloid N-oxides in Senecio vulgaris. Planta 177, 98–107.
Hartmann, T., Toppel, G., 1987. Senecionine N-oxide, the primary
product of pyrrolizidine alkaloid biosynthesis in root cultures of
The fifth youngest leaf of each plant was harvested to
Senecio vulgaris. Phytochemistry 26, 1639–1643.
Hartmann, T., Witte, L., 1995. Chemistry, biology and chemoecology
determine PA composition. The leaves were dried at
of the pyrrolizidine alkaloids. In: Pelletier, S.W. (Ed.), Alkaloids:
50 �C for 3 days and then stored at
Chemical and Biological Perspectives. Vol. 9. Pergamon Press,
were extracted by acid-base extraction (
Elmsford, New York, pp. 156–233.
PA composition of the plants was
Hartmann, T., Zimmer, M., 1986. Organ-specific distribution and
determined with GC (WCOT, 15 m�0.25 mm; DB-1
accumulation of pyrrolizidine alkaloids during the life history oftwo annual Senecio species. J. Plant Physiol. 122, 67–80.
and DB-17, J&W Scientific) with following conditions:
Macel, M., 2003. On the evolution of the diversity of pyrrolizidine
injector: 250 �C, temperature program 150–300 �C, 6 �C
alkaloids. PhD thesis. Leiden University, Leiden, The Netherlands.
per minute, carrier gas He, pressure DB-1 100 kPa and
Macel, M., Klinkhamer, P.G.L., Vrieling, K., van der Meijden, E., 2002.
M. Macel et al. / Phytochemistry 65 (2004) 865–873
Diversity of pyrrolizidine alkaloids in Senecio species does not affect
from some pyrrolizidine alkaloid producing Senecio species. Plant
the specialist herbivore Tyria jacobaeae. Oecologia 133, 541–555.
Cell. Rep 6, 466–469.
Macel, M., Vrieling, K., 2003. Pyrrolizidine alkaloids as oviposition
Vrieling, K., de Boer, N.J., 1999. Host plant choice and larval growth
stimulants for the cinnabar moth, Tyria jacobaeae. J. Chem. Ecol.
in the cinnabar moth: do pyrrolizidine alkaloids play a role? Ento-
29, 1435–1446.
mol. Exp. Appl 91, 251–257.
Mattocks, A.R., 1967. Spectrophotometric determination of unsatu-
Vrieling, K., de Vos, H., van Wijk, C.A.M., 1993. Genetic analysis of
rated pyrrolizidine alkaloids. Anal. Chem. 34, 443–447.
the concentration of pyrrolizidine alkaloids of Senecio jacobaea.
Mattocks, A.R., 1968. Toxicity of pyrrolizidine alkaloids. Nature 217,
Phytochemistry 32, 1141–1144.
Witte, L., Ernst, L., Adam, H., Hartmann, T., 1992. Chemotypes of
Toppel, G., Witte, L., Riebesehl, B., von Borstel, K., Hartmann, T.,
two pyrrolizidine alkaloid-containing Senecio species. Phytochem-
1987. Alkaloid patterns and biosynthetic capacity of root cultures
istry 31, 559–565.
Source: https://miekevd.home.xs4all.nl/PDF/81.pdf
Ginecology with ultrasound (3d) & color dopller, Mamography, Internal medicine, Pediatrics, Ortopedy CASE STUDY OF (DERMATITIS ATOPICA) TREATED WITH NEW DIETETIC PRODUCT MEGAMIN ACTIV (TMA-Z) (Preliminary results) INTRODUCTION Latin names of dermatitis atopica (AD in further text), like eczema infantum, neurodermitis, pruridermatitis allergica chronica constitutionalis Kogoj, are used. Disease with dominant recidival chronical inflamation of the skin starts usually very early, after delivery and continues in a childhood. It is usually joined with other allergic disease of child or hereditary present in a family. Neurodermitis appears in 95% of cases beore 4.th year, with first manifestations between 1-4th month of life. Clinical manifestation starts with a tiny eflorescencia like eryitema and red papulas with a strong impact of pruritus and discomfort, which leads to scratching with ecscoriation and bleeding of the skin. Tipical locations are on face, around the ears, back of head, occipital, pelvic region, extremitas (knees and elbows especially extensiors side. In some havy cases, proces can be spread over the all body. Vet manifestations on skin like vesikulas and pustulas and later appearance of crusts lead to secondary infection (bacterial, viral, rarely fungal). Manifestations on the skin show chronical progresion, lihenificated papulas mainly localised on joints, pelvic region, on neck, or diseminated. Pruritus often makes situation worse with traumatic bleeding and erosions of the skin which havilly disturb a good mud and sleeping. When signs of respiration alergy (asthma) occure, the situation becomes worse and worse. Later in adolescent faze and after, neurodermitis is present in latent form or more often recidival manifestations occure. Tipical locations of recidival dermatitis are present on flexuras or in front of lower leg or nuchal few lichenificated centers. Skin lesions can be located only on eyebrows and that could be the only clinic sign of the disease. PATOGENESIS It is not yet comletelly explained. There are few hypotesis which are trying to describe the causes of this illness. Today, one theory is mostly accepted: *allergic-immune mechanism which explains the disturbance in production of antibody (IgE, IgG4, IgA) and cell immunity. Importance of feeding in postnatal period is today of great interest in therapy, because some of the nutritive agents are considered as a 'strong alergens' like proteins of a natural milk and eggs, in relationship with the skin manifestations. Varios conservans and aditives, perorally taken drugs, inhalatory alergens are now the often subject of investigation. *Transepidermal way is also described but can not always be explained as a cause of neurodermitis because the IgE antibody can not be approved in 20-40% of cases (Buscino and all). *The other hipotesis like this which includes abnormality of beta-adrenergic receptors can not explain complete patogenesis of disease. Important role in patogenesis, which is accepted today, plays the function of Langerhans cells, which is the important factor in immune system. This cells, are members of macrofag-monocites, have the role in intracelular way of superficial antigens. Pancreatic cells carry CD1 i CD4 antigens and also Fc receptors for IgE and C3b. Beside this cells, like in a different allergic reactions, mastocites in skin release histamin which leads to eritema and pruritus (Ruzicka and all, 1983.) But, manifestations of neurodermitis can not
Sommaire 1) Ce dont traite ce document 2) Avant le départ 3) Les vaccins 4) La pharmacie du bord, un casse-tête à ranger 5) Règles simples pour éviter que les menus bobos deviennent de gros problèmes. � Repousser les microbes � Partir en balade � Chasser le hoquet � Chasser le rhume � Se prémunir contre les chocs � Aiguilles d'oursins � Piqûres d'abeilles


