
Cialis ist bekannt für seine lange Wirkdauer von bis zu 36 Stunden. Dadurch unterscheidet es sich deutlich von Viagra. Viele Schweizer vergleichen daher Preise und schauen nach Angeboten unter dem Begriff cialis generika schweiz, da Generika erschwinglicher sind.
Tumor necrosis factorrelated apoptosisinducing ligand (trail) and its death receptor (dr5) in peyronie's disease. a biomolecular study of apoptosis activation
Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand
(TRAIL) and Its Death Receptor (DR5) in Peyronie's Disease.
A Biomolecular Study of Apoptosis Activationjsm_20031.7
Carla Loreto, MD,* Guido Barbagli, MD,† Rados Djinovic, MD,‡ Giuseppe Vespasiani, MD,§Maria Luisa Carnazza, MD,* Roberto Miano, MD,§ Giuseppe Musumeci, PhD,* andSalvatore Sansalone, MD§
*Department of Anatomy, Diagnostic Pathology, Forensic Medicine, Hygiene and Public Health, University of Catania,Italy; †Centre for Reconstructive Urethral Surgery, Arezzo, Italy; ‡Serbian Academy of Science and Arts, School ofMedicine, University of Belgrade, Serbia; §Department of Urology, School of Medicine Tor Vergata University of Rome,Rome, Italy
A B S T R A C T
Introduction. Peyronie's disease (PD) is a connective tissue disorder of tunica albuginea (TA), a thick fibrous sheath
surrounding the corpora cavernosa of the penis. Relatively, little is known about the disease itself.
Aim. To investigate whether the apoptosis cascade in degenerated and macroscopically deformed TA from men with
PD is activated through the extrinsic pathway, by assessing the immunoexpression of tumor necrosis factor-related
apoptosis-inducing ligand (TRAIL) and its death receptor, DR5.
Methods. TA plaques from 15 men with PD and from four unaffected men were processed for TRAIL and DR5
immunohistochemistry and Western blot analysis.
Main Outcome Measures. A greater understanding of the pathophysiology of PD through a molecular approach, to
gain insights that may lead to novel forms of treatment.
Results. Activation of the apoptosis mechanisms through the extrinsic pathway was demonstrated by TRAIL and
DR5 overexpression in fibroblasts and myofibroblasts from affected TA.
Conclusion. The finding that apoptosis activation in TA plaques occurs, at least in part, via the extrinsic pathway may
help devise novel therapeutic options for these patients.
Loreto C, Barbagli G, Djinovic R, Vespasiani G,
Carnazza ML, Miano R, Musumeci G, Sansalone S. Tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL) and its death receptor (DR5) in Peyronie's disease. A biomolecular study of apoptosis
activation. J Sex Med **;**:**–**.
Key Words. Peyronie's Disease; Peyronie's Plaque Healing; Apoptosis; TRAIL; DR5
sexual activity [7]. There is growing consensusthat the resulting TA injury or tear heals abnor-
P eyronie's disease (PD) is a connective tissue mally, although the underlying mechanism is
disorder of the tunica albuginea (TA), a thick
unclear [6,8–11]. Some researchers view PD
fibrous sheath surrounding the corpora cavernosa
plaques as scars that have failed to remodel well
of the penis [1–4]. It affects 2–3% of the male
[11]. Relatively little is known about the disease
population between the 4th and the 6th decade
itself; this is also reflected in the lack of effective
[5,6] and is characterized clinically by plaques,
treatments capable of altering its course or pro-
penis deformation, localized pain, and erectile
gression [7,12]. A greater understanding of the
dysfunction. The initiating event in plaque devel-
pathophysiology of PD at the molecular level
opment seems to be an external stress received
may provide insights and lead to novel forms of
most likely in the erect state, usually during
2010 International Society for Sexual Medicine
J Sex Med **;**:**–**
Loreto et al.
The working hypothesis of this study was that
after intracavernous injection of alprostadil
apoptosis, or programmed cell death, may be
implicated in PD. Apoptosis is a tightly regulated
The control samples were from four patients
process involved in embryogenesis, metamorpho-
(mean age 23 ⫾ 3.0 years; range 21–27) with con-
sis, normal tissue turnover, wound healing, and a
genital penile curvature, who underwent Nesbit's
variety of pathological conditions resulting from
corrective procedure [17]. Their clinical histories
its dysregulation, whose function is to remove
were negative for generalized penile diseases
harmful, damaged, or unwanted cells [13]. This
and none had macroscopic signs of degenerative
homeostatic mechanism acts through two main
or inflammatory disorders. Staining of these
pathways: the extrinsic (or death receptor-
specimens with Mayer's hematoxylin showed no
mediated) and the intrinsic (or mitochondrial).
detectable pathological abnormalities on light
The former involves activation of death signaling
ligands, like tumor necrosis factor (TNF)-a, FasL,tumor necrosis factor-related apoptosis-inducingligand (TRAIL; a recently identified death factor
that acts as a potent apoptosis inducer and activates
For the immunohistochemical studies, the TA
its death receptors DR4 and DR5), and members
samples were fixed overnight in 10% neutral buff-
of the TNF-receptor superfamily (e.g., TNF
ered formalin (Bio-Optica, Milan, Italy) and then
receptor I, DR4, and DR5), which results in acti-
demineralized in ethylenediaminetetraacetic acid
vation of caspase 8, that in turn cleaves—hence
(EDTA) decalcification fluid (41.3 g disodium
activates—the proenzyme form of caspase 3, an
EDTA, 4.4 g NaOH in 1,000 mL distilled water)
executioner caspase. The latter enzyme then per-
for 6 weeks at 4°C. After fixation and overnight
forms its executioner role by destroying the cell's
washing, they were dehydrated in graded ethanol
cytoskeletal and reparative proteins [14,15], a
and embedded in paraffin. Specimens were then
process that culminates with DNA fragmentation.
sectioned at a thickness of 5 mm and placed on
We investigated whether the extrinsic pathway
silanized glass slides. Endogenous peroxidase
participates in apoptosis activation by assessing the
activity was quenched by treatment with 3%
immunoexpression of TRAIL and its death recep-
H2O2 for 10 minutes. Nonspecific antibody
tor, DR5, in TA plaques to gain insights into the
binding was blocked by treatment with normal
molecular activation of programmed cell death in
horse/goat serum (dil. 1:20 in phosphate buffered
PD. To our knowledge, there are no similar animal
saline, 0.1% bovine serum albumin). Sections were
or human studies.
irradiated (5 minutes ¥ 3) in capped polypropyleneslide-holders with citrate buffer (pH 6), using amicrowave oven (750 W) to unmask antigen sites.
Materials and Methods
Antibodies for localization of TRAIL and DR5
Patients and Tissues
were rabbit polyclonal anti-TRAIL (Santa Cruz
Biopsy specimens (wedge shaped, approximately
Biotechnology, Inc., Santa Cruz, CA, USA) and
5 ¥ 3 mm) were collected from 15 patients (mean
anti-DR5 (Novus Biologicals, Littleton, CO,
age 53 ⫾ 10 years; range 31–67) with stable PD for
USA) used at 1:100 working dilutions. They were
at least 6 months at the level of the corporotomy
applied directly onto sections and slides were incu-
during corrective surgery for PD. All patients
bated overnight (4°C) in a humid chamber.
underwent albugineal grating using the geometri-
Immune complexes were then treated with a bioti-
cal principle, as originally described by Egydio
nylated link antibody and then detected with
[16]. The TA was incised and grafted at the level of
peroxidase-labeled streptavin, both incubated for
the maximum curvature, where plaque was most
10 minutes at room temperature (LSAB + System-
HRP, Dako Italia SPA, Milan, Italy).
The study was approved by the local ethics
The immunoreaction was examined with an
committee. The informed consent of each patient
Axioplan light microscope (Zeiss, Oberkochen,
was obtained before tissue collection.
Germany) after incubating sections in 0.1% 3,3′-
On preoperative examination, all patients
diaminobenzidine and 0.02% hydrogen peroxide
reported spontaneous erections, but being pre-
solution (DAB substrate kit, Vector Laboratories,
vented from having regular sexual intercourse by a
Burlingame, CA, USA) for 4 minutes. Sections
penile curvature >45°. Degree of curvature and
were lightly counterstained with Mayer's hema-
rigidity were evaluated with Doppler ultrasound
J Sex Med **;**:**–**
Apoptosis in Peyronie's Disease
Sweden) and finally mounted on GVA mount
and with a secondary peroxidase-conjugated anti-
(Zymed Laboratories, San Francisco, CA, USA).
rabbit (Amersham Pharmacia Biotech; 1:10,000)antibody or a polyclonal rabbit anti-DR5 antibody
(Santa Cruz Biotechnology). Detection was per-
The intensity of TRAIL and DR5 staining and the
formed with a chemiluminescence assay (ECL;
proportion of immunopositive cells were exam-
Amersham Italia, Milano, Italy).
ined in blind by light microscopy and recorded by
All experiments were performed at least three
two anatomists and a histologist. Intensity of stain-
times and the signal intensity was analyzed using a
ing (IS) was graded in a semiquantitative manner
digital imaging analysis system (1D Image Analysis
using a 5-point scale, as follows: 0 = no detectable
Software; Scientific Imaging Kodak, New Haven,
staining, 1 = weak staining, 2 = moderate staining,
CT, USA). b-tubulin (Santa Cruz Biotechnology)
3 = strong staining, 4 = very strong staining. The
was used as an internal control to validate the right
proportion of TRAIL- and DR5-immunopositive
amount of protein loaded onto the gels.
cells (extent score = ES) was evaluated indepen-
Densitometric analysis was expressed as inte-
dently by two anatomists and a histologist and
grated density by normalizing sample values to the
scored as a proportion of 200 cells into four cat-
corresponding b-tubulin expression.
3 = >50%, and 4 = >75%. Counting was per-formed at 200¥ magnification. The final staining
score (FSS) was the sum of IS + ES.
Mean and standard deviation (SD) were calculatedfor the FSS. All data were analyzed with the SPSS
Positive and Negative Controls
program (SPSS® release 16.0, Chicago, IL, USA).
Positive and negative controls were performed totest the specific reaction of primary antibodies atthe protein level. For positive control testing, basal
cell carcinoma tissue was exposed to an immunop-eroxidase process. Immunolabeling for TRAIL
Histological examination of hematoxylin-stained
and DR5 was found both in membranes and cyto-
specimens showed that the collagen fiber arrange-
plasm. For negative controls, TA plaques were
ment was affected to different extents in all PD
treated with normal rabbit serum instead of the
samples, with damage ranging from fiber fragmen-
tation to tears and splitting. The normal collagenfiber arrangement and ordered appearance was
predominantly lost. The elastic fibers were often
Fresh TA tissue was homogenized in a Polytron
fragmented; in some cases, the collagen bundles
homogenizer using a lysis buffer containing
formed clumps with surrounding elastic fibers.
150 mM NaCl, 50 mM Tris–HCl (pH 7.5), 5 mM
In contrast, the control TA samples showed
EDTA, 1 mM Na3VO4, 30 mM Na pyrophos-
preserved collagen bundles, with longitudinal fiber
phate, 50 mM NaF, 1 mM acid phenyl-methyl-
orientation in the outer layer and circular bundles
sulfonyl-fluoride, 5 mg/mL aprotinin, 2 mg/mL
in the inner layer.
leupeptin, 1 mg/mL pepstatin, 10% glycerol, and
Immunohistochemical examination of PD sec-
0.2% Triton X-100. The homogenates were then
tions showed immunoexpression of both TRAIL
centrifuged at 14,000 rpm for 10 minutes at 4°C.
and DR5 in fibroblasts and myofibroblasts (2–3),
The protein concentration of the supernatant was
respectively, in the cytoplasm and in membrane
determined by the Bradford method [18].
and cytoplasm (Figures 1 and 2), whereas few orno immunostained cells were observed in control
Western Blot Analysis
samples (1) (Figure 3). The proportion of immu-
Equal amounts of protein were subjected to
nopositive cells and the intensity of the immunore-
sodium dodecyl sulfate–polyacrylamide gel elec-
action in PD patients were much higher than in
trophoresis on 10% gels, transferred onto Hybond
normal tissue. The mean FSS of TA plaques was
ECL nitrocellulose membranes (Amersham Phar-
4.77 ⫾ 0.61 (range 3.6–5.6).
macia Biotech, Amersham, UK) for 1 hour, and
Western blot analysis of PD samples showed
analyzed by immunoblotting with a primary poly-
significantly increased TRAIL expression associ-
clonal anti-TRAIL antibody (BD Transductions
ated with constitutive expression of DR5 receptor
Laboratories, Franklin Lakes, NJ, USA; 1:1.000),
protein (Figure 4).
J Sex Med **;**:**–**
Loreto et al.
Figure 1 TRAIL immunostaining in a
tunica albuginea plaque from a subject
with Peyronie's disease (A) (¥40). (B)
Magnification of the inset in (A):
TRAIL-positive fibroblast and myofi-
broblast cells.
Using a TGF-b1-injected rat model, El-Sakka
and colleagues documented an increase in myofi-
To the best of our knowledge, this is the first study
broblast content, endogenous TGF-b1 expression
addressing some molecules critically implicated in
and collagen synthesis in diseased TA [22–27]
the TA apoptotic cascade in PD patients. In this
similar to those seen in human PD. These charac-
preliminary investigation, we focused on the acti-
teristics are accompanied by an increase in reactive
vation of the extrinsic pathway and DR5 expres-
oxygen species (ROS), a well-known profibrotic
sion. The disease is characterized by localized TA
factor, and in inducible nitric oxide synthase
disruption, a local increase in microvascular per-
(iNOS), a recently recognized antifibrotic com-
meability, persistent fibrin (deficient fibrinolysis)
pound, whose induction may arise as a defense
and collagen buildup, perivascular inflammation,
mechanism against oxidative stress and fibrosis.
disruption and loss of elastic fibers, disorganized
The iNOS nitric oxide (NO) quenches ROS by
collagen bundles, increased synthesis of trans-
forming peroxynitrite, an inducer of fibroblast and
forming growth factor b1 (TGF-b1), and cell- and
myofibroblast apoptosis [24–26,28,29].
collagen fiber-mediated contraction of pathologi-
TGF-b1 is a multifunctional cytokine that
cal TA, with eventual calcification and ossification
inhibits growth and induces apoptosis of rat liver
[1–4,8–11,19–21]. Scanty data are available on the
epithelial cells [30]; apoptosis is blocked if TGF-b1
molecular mechanisms underlying PD.
expression is downregulated [31]. TGF-b1 can also
Figure 2 DR5 immunostaining in a
tunica albuginea plaque from a subject
with Peyronie's disease (A) (¥40). (B)
Magnification of the inset in (A): DR5-
positive fibroblast and myofibroblast
cells.
Figure 3 Immunostaining for TRAIL
(A) and its receptor DR5 (B) in control
samples of tunica albuginea from a
subject without Peyronie's disease
(¥20).
J Sex Med **;**:**–**
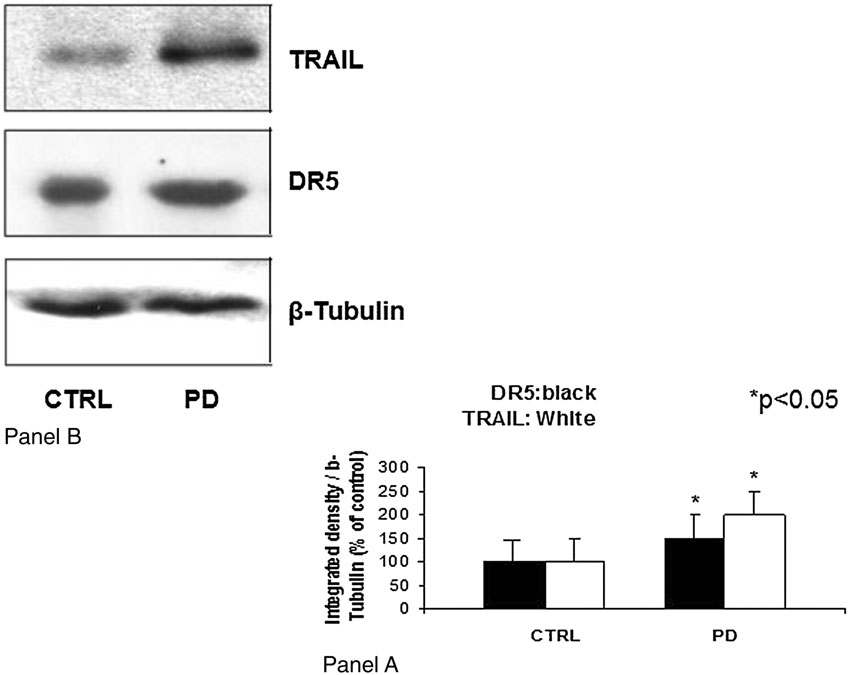
Apoptosis in Peyronie's Disease
Figure 4 Expression of TRAIL and its
death receptor DR5 in TA plaques from
a patient with Peyronie's disease.
Panel A: integrated density of the
Western
*P < 0.05 vs.
control values; Panel B: Western blotanalysis of proteins in TA plaques froma patient with Peyronie's disease.
Upper blot: TRAIL expression; middleblot: expression of DR5 death recep-tor;
control; PD: Peyronie's disease.
influence fibroblast survival and induce fibrosis in
and the halting of fibrosis progression. However, it
chronic inflammatory disorders [31–33].
should be considered that after its activation, the
TRAIL is a member of the TNF family of
apoptosis process could be blocked by endogenous
ligands, capable of initiating apoptosis through
apoptosis inhibitors. Further studies are therefore
engagement of its death receptors (DR4, DR5). It
needed to gain insights into this delicate process.
selectively induces apoptosis of a variety of tumor
Apoptosis of TA cells may also be triggered by
and transformed cells, but not of most normal cells
increased tissue tension and compression, as dem-
[34], and has been seen to play a large role in cell
onstrated in vitro in intervertebral discs, where it
regulation and in inflammation processes [35,36].
has been suggested that a threshold of tissue strain
Overall, four different receptors have been identi-
initiates programmed cell death in the anulus
fied that interact with TRAIL, the two death-
fibrosus [42]; it has also been demonstrated that
inducing receptors, DR4 and DR5, and DcR1 and
myofibroblasts are more than likely to play a
DcR2 (decoy receptors), which do not induce
central role in mechanotransduction [43,44]. Con-
death upon ligation and are believed to counteract
sidering that cells are likely to play a substantial
TRAIL-induced cytotoxicity [37]. DR5 is a widely
role in tissue turnover, and that tissue degenera-
investigated apoptosis molecule, also in a number
tion seems to be associated to a loss of cellularity
of tissue diseases that correlate with degenerative
through apoptosis-related processes, the latter
process may lead to decreased extracellular matrix
generation and affect tissue organization and
myofibroblast proliferation and accumulation of
repair mechanisms [38]. In this regard, we demon-
extracellular matrix [41]. In particular, TGF-b1
strated through TRAIL and DR5 upregulation
promotes a fibroblast-to-myofibroblast transition
that apoptosis activation could be responsible for
that is initially followed by myofibroblast replica-
tissue degeneration besides being a defense
tion and activation, leading to an excess of collagen
synthesis. In normal healing, a protective mecha-
Despite a well-recognized effect on the TA, age
nism achieves myofibroblast clearance through the
does not seem to influence the tissue expression of
apoptosis process. If the mechanism fails, persis-
the apoptosis molecules in a significant way, as
tence of these cells induces fibrosis with collagen
inferred from studies of intervertebral and tem-
accumulation and tissue contraction [7]. In this
poromandibular disc programmed cell death
regard, we documented activation of the mecha-
involving experimental and control groups with
nism through overexpression of DR5 and its
very different mean ages [38,45] or describing a
ligand, which could underpin plaque stabilization
lack of statistical correlation with age [46].
J Sex Med **;**:**–**
Loreto et al.
This work documented activation of apoptosis
(b) Revising It for Intellectual Content
through the extrinsic pathway in fibroblasts and
Guido Barbagli; Rados Djinovic
myofibroblasts from PD patients as overexpressionof TRAIL and its death receptor, DR5. We intend
Category 3
to follow up on these findings by exploring other
(a) Final Approval of the Completed Manuscript
molecules involved in programmed cell death.
Carla Loreto; Salvatore Sansalone
Further research efforts could be extended toaddress the intrinsic pathways and executive
caspases. We are also interested in investigatingthe action of TRAIL on fibroblast cell lines and on
1 Jordan GH, Schlossberg SM, Devine CJ. Surgery of the penis
and urethra. In: Walsh PC, Retik AB, Vaughan ED, Wein AJ,
isolated plaque-derived human myofibroblasts
eds. Campbell's urology. 7th edition. Philadelphia: WB Saun-
[47]. These further studies have the potential to
2 Noss MB, Day NS, Christ GJ, Melman A. The genetics and
involved in PD. The study of apoptosis could
immunology of Peyronie's disease. Int J Impot Res 2000;4:S127–390.
provide a better understanding of the underlying
3 Hellstrom WJ, Bivalacqua TJ. Peyronie's disease: Etiology,
mechanisms of this complex process, to devise
medical, and surgical therapy. J Androl 2000;21:347–54.
potential therapeutic strategies, for instance, based
4 Gholami SS, Lue TF. Peyronie's disease. Urol Clin North Am
on TRAIL agonists. In PD patients, this could be
5 Rhoden EL, Teloken C, Ting HY, Lucas ML, Teodósio da Ros
through enhancement of myofibroblast death in
C, Ary Vargas Souto C. Prevalence of Peyronie's disease in
the attempt to induce plaque stabilization and
men over 50 years old. J Urol 2001;165:200.
prevent disease progression.
6 McVary KT. Peyronie's disease. Paper presented at the annual
meeting of the American Urological Association, Anaheim,CA, 2001.
7 Gonzalez-Cadavid NF, Magee TR, Ferrini M, Qian A, Vernet
D, Rajfer J. Gene expression in Peyronie's disease. Int J Impot
In conclusion, apoptotic cell death in TA plaques
occurs, at least in part, via the extrinsic pathway.
8 Davis CJ. The microscopic pathology of Peyronie's disease.
This finding could have important therapeutic
J Urol 1997;157:282–4.
9 Devine CJ, Somers KD, Jordan SG, Schlossberg SM. Pro-
posal: Trauma as the cause of the Peyronie's lesion. J Urol1997;157:285–90.
10 Jarow JP, Lowe FC. Penile trauma: An etiologic factor in
Peyronie's disease and erectile dysfunction. J Urol 1997;158:
The authors wish to remember their late colleague Pro-
fessor S. Perovic, who worked with them on this project
11 Ehrlich HP. Scar contracture: Cellular and connective tissue
aspects in Peyronie's disease. J Urol 1997;157:316–9.
before his premature death.
12 Hellstrom WJG, Cockerham YR. Intralesional injection
therapy for Peyronie's disease. J Sex Med 2009;6:1811–4.
Corresponding Author: Carla Loreto, MD, Depart-
13 Huppertz B, Kaufmann HGFP. The apoptosis cascade-
ment of Anatomy, Diagnostic Pathology, Forensic
morphological and immunohistochemical methods for its
Medicine, Hygiene and Public Health, University of
visualization. Anat Embryol 1999;200:1–18.
Catania, Via S. Sofia, 87-95123, Catania 95123, Italy.
14 Tschoeke SK, Hellmuth M, Hostmann A, Robinson Y, Ertel
Tel: 390953782038; Fax: 390953782046; E-mail:
W, Oberholzer A, Heyde CE. Apoptosis of human interverte-
bral discs after trauma compares to degenerated discs involvingboth receptor-mediated and mitochondrial-dependent path-
Conflict of Interest: None.
ways. J Orthop Res 2008;26:999–1006.
15 Ferri KF, Kroemer G. Organelle-specific initiation of cell
death pathway. Nat Cell Biol 2001;3:255–63.
Statement of Authorship
16 Egydio PH, Sansalone S. Peyronie's reconstruction for
maximum length and girth gain: Geometrical principles. Adv
Category 1
Urol 2008;2008:205739 (Epub 2008).
(a) Conception and Design
17 Nesbit RM. Congenital curvature of the phallus: Report of
Carla Loreto; Salvatore Sansalone
three cases with description of corrective operation. J Urol
(b) Acquisition of Data
18 Bradford MM. A rapid and sensitive method for the quan-
Giuseppe Musumeci; Roberto Miano
titation of microgram quantities of protein utilizing the
(c) Analysis and Interpretation of Data
principle of protein-dye binding. Anal Biochem 1976;72:
Maria Luisa Carnazza; Giuseppe Vespasiani
19 Somers KD, Dawson DM. Fibrin deposition in Peyronie's
Category 2
disease plaque. J Urol 1997;157:306–16.
20 Shindel AW, Lin G, Ning H, Banie L, Huang YC, Lin G, Lin
(a) Drafting the Article
CS, Lue FL. Pentoxifylline attenuates transforming growth
Carla Loreto; Salvatore Sansalone
factor-ß1-stimulated collagen deposition and elastogenesis in
J Sex Med **;**:**–**
Apoptosis in Peyronie's Disease
human tunica albuginea-derived fibroblasts part 1: Impact on
35 Hasel C, Durr S, Rau B, Strater J, Schmid RM, Walczak H,
extracellular matrix. J Sex Med 2010;7:1787–97.
Bachem MG, Moller P. In chronic pancreatitis, widespread
21 Young LX, Bella AJ, O'Gorman DB, Gan BS, Lim KB, Brock
emergence of TRAIL receptors in epithelia coincides with
GB. Protein biomarker analysis of primary Peyronie's disease
neoexpression of TRAIL by pancreatic stellate cells of early
cells. J Sex Med 2010;7:99–106.
fibrotic areas. Lab Invest 2003;83:825–36.
22 El-Sakka AL, Hassoba HM, Chui RM, Bhatnagar RS,
36 Robertson NM, Rosemiller M, Lindemeyer RG, Steplewski A,
Dahiya R, Lue TF. An animal model of Peyronie's-like
Zangrilli JG, Litwack G. TRAIL in the airways. Vitam Horm
condition associated with an increase of trasforming growth
factor beta mRNA and protein expression. J Urol 1997;158:
37 Sanlioglu AD, Korcum AF, Pesterelli E, Erdogan G, Karaveli
S, Savas B, Griffith TS, Sanlioglu VMD. Trail death
23 El-Sakka AL, Hassan MU, Nunes L, Bhatnagar RS, Yen TS,
receptor-4 expression positively correlates with the tumor
Lue TF. Histological and ultrastructural alterations in an
grade in breast cancer patients with invasive ductal carcinoma.
animal model of Peyronie's disease. Br J Urol 1998;81:445–52.
Int J Radiat Oncol Biol Phys 2007;69:716–23.
24 Ferrini MG, Vernet D, Magee TR, Shahed A, Qian A, Rajfer
38 Leonardi R, Almeida LE, Trevilatto PC, Loreto C. Occur-
J, Gonzalez-Cadavid NF. Antifibrotic role of inducible nitric
rence and regional distribution of TRAIL and DR5 on tem-
oxide synthase. Nitric Oxide 2002;6:283–94.
poromandibular joint discs: Comparison of disc derangement
25 Ferrini MG, Magee TR, Vernet D, Rajfer J, Gonzalez-Cadavid
with and without reduction. Oral Surg Oral Med Oral Pathol
NF. Aging-related expression of inducible nitric oxide syn-
Oral Radiol Endod 2010;109:244–51.
thase and markers of tissue damage in rat penis. Biol Reprod
39 Loreto C, Musumeci G, Leonardi R. Chondrocyte-like apop-
tosis in temporomandibular joint disc internal derangement as
26 Vernet D, Ferrini MG, Valente EG, Magee TR, Bou-Gharios
a repair-limiting mechanism. An in vivo study. Histol Histo-
G, Rajfer J, Gonzalez-Cadavid NF. Effect of nitric oxide on
the differentiation of fibroblasts into myofibroblasts in the
40 Cui M, Wang L, Liang X, Ma X, Liu Y, Yang M, Liu K,
Peyronie's fibrotic plaque and in its rat model. Nitric Oxide
Wei X, Zhou Z, Chen YH, Sun W. Blocking TRAIL-DR5
signaling with soluble DR5 reduces delayed neuronal damage
27 Gonzalez-Cadavid NF, Rajfer J. Experimental models of
after transient global cerebral ischemia. Neurobiol Dis
Peyronie's disease. Implication for new therapies. J Sex Med
41 Pardo A, Barrios R, Maldonado V, Meléndez J, Ruiz V, Segura
28 Bivalacqua TJ, Diner EK, Novak TE, Vohra Y, Sikka SC,
L, Sznajder JI, Selman M. Gelatinases A and B are upregulated
Champion HC, Kadowitz PJ, Hellstrom WJ. A rat model of
in lung rats by subacute hyperoxia. Pathogenetic implications.
Peyronie's disease associated with a decrease in erectile activity
Am J Pathol 1998;153:833–44.
and an increase in inducible nitric oxide synthase protein
42 Setton LA, Chen J. Mechanobiology of the intervertebral disc
expression. J Urol 2000;163:1992–8.
and relevance to disc degeneration. J Bone Joint Surg Am
29 Davila HH, Ferrini MG, Rajfer J, Gonzalez-Cadavid NF.
Fibrin as an inducer of fibrosis in the tunica albuginea of the
43 Wang J, Zohar R, McCulloch CA. Multiple roles of alpha
rat: A new animal model of Peyronie's disease. BJU Int
smooth muscle actin in mechanotransduction. Exp Cell Res
30 Teramoto T, Kiss A, Thorgeirsson SS. Induction of p53 and
44 Gelbard M. Myofibroblasts and mechanotransduction: Do
Bax during TGF-beta 1 initiated apoptosis in rat liver epithe-
forces in the tunica albuginea contribute to Peyronie's disease?
lial cells. Biochem Biophys Res Commun 1998;251:56–60.
J Sex Med 2008;5:2973–6.
31 Sulkowski S, Wincewicz A, Sulkowska M, Koda M. Trans-
45 Chen B, Ma B, Yang S, Xing X, Gu R, Hu Y. DR5 and DcR2
forming growth factor-beta1 and regulators of apoptosis. Ann
are expressed in human lumbar intervertebral discs. Spine
N Y Acad Sci 2009;1171:116–23.
32 Mulsow JJ, Watson RW, Fitzpatrick JM, O'Connell PR. Trans-
46 Gruber HE, Hanley EN. Analysis of aging and degeneration of
forming growth factor-beta promotes pro-fibrotic behavior by
the human intervertebral disc: Comparison of surgical speci-
serosal fibroblasts via PKC and ERK1/2 mitogen activated
mens with normal control. Spine 1998;23:751–7.
protein kinase cell signaling. Ann Surg 2005;242:880–7.
47 Piao SP, Choi MJ, Tumurbaatar M, Kim WJ, Jin HR, Shin
33 McKaig BC, Hughes K, Tighe PJ, Mahida YR. Differential
SH, Tuvshintur B, Yin GN, Song JS, Kwon MH, Lee SJ, Han
expression of TGF-beta isoforms by normal and inflammatory
JY, Kim SJ, Ryu JK, Suh JK. Trasforming growth factor
bowel disease intestinal myofibroblasts. Am J Physiol Cell
(TGF)-ß type I receptor kinase (ALK5) inhibitor alleviates
profibrotic TGF-ß1 responses in fibroblasts derived from
34 Wang S, El-Deiry WS. TRAIL and apoptosis induction by
Peyronie's plaque. J Sex Med 2010; Mar 11 [Epub ahead of
TNF-family death receptors. Oncogene 2003;22:8628–33.
J Sex Med **;**:**–**
Source: http://www.peyroniecenter.it/files/Tumor%20Necrosis%20Factor-Related%20Apoptosis-Inducing%20Ligand.pdf
Author's personal copy Food Microbiology 28 (2011) 214e220 Contents lists available at ScienceDirect Food Microbiology Bacillus probiotics Simon M. Cutting* School of Biological Sciences, Royal Holloway University of London, Egham, Surrey TW20 0EX, UK Bacterial spore formers are being used as probiotic supplements for use in animal feeds, for human Available online 24 March 2010
MATERIAL SAFETY DATA SHEET Page 1 of 8 MSDS-120 Prepared to OSHA, ACC, ANSI, NOHSC, WHMIS & 2001/58 EC Standards MSDS Revision: 1.0 MSDS Revision Date: 06/14/2010 1. PRODUCT IDENTIFICATION ALL ACRYLIC POLYMER POWDERS - All Shades ACRYLATES COPOLYMER POWDER PROFESSIONAL OR SUNDRY USE ONLY


