
Cialis ist bekannt für seine lange Wirkdauer von bis zu 36 Stunden. Dadurch unterscheidet es sich deutlich von Viagra. Viele Schweizer vergleichen daher Preise und schauen nach Angeboten unter dem Begriff cialis generika schweiz, da Generika erschwinglicher sind.
Npgrj_ng_1616 937.944
Contributions of low molecule number and
enetics chromosomal positioning to stochastic gene expression
Attila Becskei1, Benjamin B Kaufmann1,2 & Alexander van Oudenaarden1
The presence of low-copy-number regulators and switch-like signal propagation in regulatory networks are expected to increasenoise in cellular processes. We developed a noise amplifier that detects fluctuations in the level of low-abundance mRNAs in
.nature yeast. The observed fluctuations are not due to the low number of molecules expressed from a gene per se but originate in the
random, rare events of gene activation. The frequency of these events and the correlation between stochastic expressions of genesin a single cell depend on the positioning of the genes along the chromosomes. Transcriptional regulators produced by suchrandom expression propagate noise to their target genes.
In living cells, fluctuations of molecule copy number are inevitable
noise in regulatory genes is believed to be determined mostly by
under certain conditions. On one hand, such random fluctuations
mRNA rather than protein levels13.
may impair signal propagation and hamper the coordination of
In contrast to intrinsic noise, extrinsic noise does not depend on the
lishing Gr
cellular activities1. On the other hand, noise in gene expression
number of molecules expressed from a gene but stems from fluctua-
introduces phenotypic heterogeneity in an isogenic population,
tions in the transcription factor activity that regulates the gene. Such
which may facilitate cellular differentiation or may be beneficial in
fluctuations might arise from environmental heterogeneity or from
temporally or spatially heterogeneous environments2,3.
global cell-to-cell variations in metabolic or biosynthetic activities8.
Total noise is typically divided into two components: intrinsic and
These fluctuations, even if small, can be magnified when they are
extrinsic4,5. Intrinsic noise, by definition, originates in the randomness
coamplified along with signals during signal amplification. Amplifica-
2005 Nature Pub associated with discrete, rare biomolecular events (e.g., mRNA synth- tion is crucial, for example, in linear transcriptional networks where
esis), when few molecules are involved6,7. The remaining noise, which oscillatory signals of the cell cycle are considerably damped during
measures fluctuations in the regulation of a gene, is lumped together
their propagation14. Feedforward and feedback loops, and regulatory
as extrinsic noise6. Noise passed on from upstream transcription
switches generated by multistep phosphorylation or cooperative
factors, as well as changes in the global and local cellular environment
binding, are capable of counteracting the dampening of signals15–19.
of a gene, can all contribute to this component8,9.
In addition to amplifying the signal, transcription factors acting
When the number of molecules is low, the discrete nature of
through positive feedback or ultrasensitive switches can generate
molecular concentrations becomes pronounced. If the steady-state
considerable cell-to-cell variation in gene expression2,20–22.
concentration corresponds to only a few molecules per cell, changing
Given that a large proportion of mRNAs is expressed at low level,
the concentration of a molecular species by even a single molecule is
noise due to the low number of molecules is expected to be great. How
relatively substantial. Protein concentration ranges from 50 to one
intrinsic noise affects regulators expressed at very low levels has not
million copies per cell in yeast10, and mRNA abundance varies
been explored. Noise in gene expression is usually measured by the
between 0.001 and 100 copies per cell. Fully 75% of yeast genes
cell-to-cell variation in expression of fluorescent reporters driven by
have steady-state transcript levels of one or fewer copies per cell
promoters of interest23,24. Intrinsic and extrinsic noise in gene
averaged over a cell population11,12. mRNAs encoding transcription
expression can be quantified by monitoring the expression of two
factors, including those regulating the cell cycle, are usually expressed
fluorescent proteins of different colors driven by the same promoter6.
at low to very low levels. For example, the steady-state level of mRNAs
In this case, uncorrelated fluctuations in reporter gene expression
encoding the cell cycle transcriptional regulators SWI6, SWI4 and
reflect intrinsic noise, because the realization of rare probabilistic
SWI5 have been reported to be 0.005, 0.3 and 1 copy per cell,
events of mRNA production is not correlated between the two genes.
respectively11. Among all expressed genes, SWI6 belongs to the set
Extrinsic noise is usually associated with correlated fluctuations,
of genes expressed at the lowest level. Because the copy number of
because fluctuations in the concentration of a transcription factor
mRNAs is typically small compared with protein number, intrinsic
are transmitted equally to the identical promoters.
1Department of Physics, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139, USA. 2Division of Engineering and Applied Sciences, HarvardUniversity, Cambridge, Massachusetts 02138, USA. Correspondence should be addressed to A.v.O. (
[email protected]).
Published online 7 August 2005; doi:10.1038/ng1616
NATURE GENETICS VOLUME 37 [ NUMBER 9 [ SEPTEMBER 2005
When fluorescent proteins are expressed at very low levels, cellular
YFP fluorescence intensity. The ratio of x2 to x1 can be varied by
background fluorescence hampers direct noise measurement by the
adjusting the doxycycline concentration. Thus, the input signals
two-color method. To circumvent this limitation, we developed an
transmitted from both weak and strong promoters can be tuned to
amplifier of fluctuations in gene expression. We then raised the
produce the same mean output signal while the corresponding noise
number of a given mRNA molecule by multiplying the number of
intensities reflect faithfully the difference in the input noise. This
gene copies encoding the mRNA species. Increased molecule number
method, therefore, offers the advantage to measure different noise
is expected to reduce the intrinsic noise. In this work, we calculate the
levels at same intensity of fluorescence. The output noise Z2 is
contribution of intrinsic noise to total noise from the ratio of
calculated by dividing the standard deviation of the YFP expression
amplified noise intensity of a gene having multiple copies to that of
distribution in a cell population by the mean YFP expression (Fig. 1b);
a single-copy gene in the genome.
this is also called the coefficient of variation. To determine the value ofthe input noise, noise transmitted to the output module must be
measured accurately. For this purpose, we explored how the input
Design of a genetic circuit to amplify fluctuations
noise can be amplified.
To amplify the signal generated by weak promoters, we designed a
Noise in the output module Z2 is directly proportional to the
synthetic gene network in the yeast Saccharomyces cerevisiae (Fig. 1a).
product of noise transmitted from the input module Z1 and the
A yeast promoter of interest drives the expression of a potent
logarithmic gain H21 of the promoter when intrinsic noise due to low
.com/natureg transcriptional activator, rtTA, in the input module. When rtTA is
copy numbers of YFP mRNA or protein is negligible. H21 reflects how
bound to the inducer, doxycycline, it drives the expression of yellow
a relative change in the input signal affects the output signal (Fig. 1a)7.
fluorescent protein (YFP) in the response module. Thus, the input
A theoretical analysis shows that increasing the cooperativity of rtTA
signal (expression strength of rtTA, x1) of the yeast promoter is
binding to the response module should amplify fluctuations. Assum-
amplified, and the resulting output signal x2 is measured by the
ing that a Hill-type function describes the cooperative binding of rtTAto the response promoter leads to a simple direct proportionality
between H21 and the Hill coefficient n. Thus, a promoter with a Hillcoefficient of 2 doubles the output noise Z2 relative to a noncoopera-
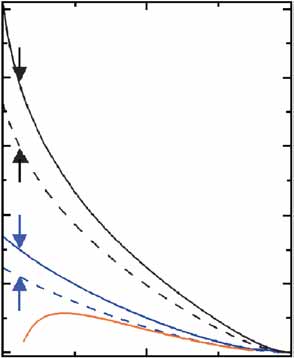
tive (n ¼ 1) but otherwise identical promoter (Fig. 1c). Z2 decreases
monotonously as the output signal x2 increases, unless basal tran-scription x
2,bas sets the lower limit of promoter activity (Fig. 1c). In
the latter case, H
21 reaches a maximum when the output signal
lishing Gr
x2;basx2;max and vanishes when the output signal approaches
either the basal expression x2,bas or the maximal expression level x2,max
(Fig. 1c and Supplementary Note online).
We used the inherently cooperative binding of transcription factors
to multiple binding sites around chromatin to increase cooperativity.
The first binding event alters chromatin structure, which enhances the
binding of the next transcription factor to the neighboring binding
2005 Nature Pub
sites25,26. This chromatin-mediated cooperativity occurs even in the
absence of direct protein-protein interaction between transcription
2 output noise ≈ H21 1
factors. To test the effect of cooperativity, response promoters contain-
ing one, two or seven binding sites (PTETO1, PTETO2 or PTETO7) for
rtTA were incorporated in the synthetic network (Fig. 1a).
�2 = x
Cooperativity-based amplification of fluctuations
Varying the concentration of doxycycline generates a response func-
tion characteristic of the promoter in the output module (Fig. 2a). We
used the Hill coefficient as a measure of sensitivity (switch-like nature)of promoter response. When the input module contained the SWI5
promoter, PSWI5, the Hill numbers were 0.95, 1.1 and 1.45 for PTETO1,
Single-cell fluorescence
Output signal x2
PTETO2 and PTETO7, respectively. Therefore, the sensitivity of theresponse increased with but was not proportional to the number of
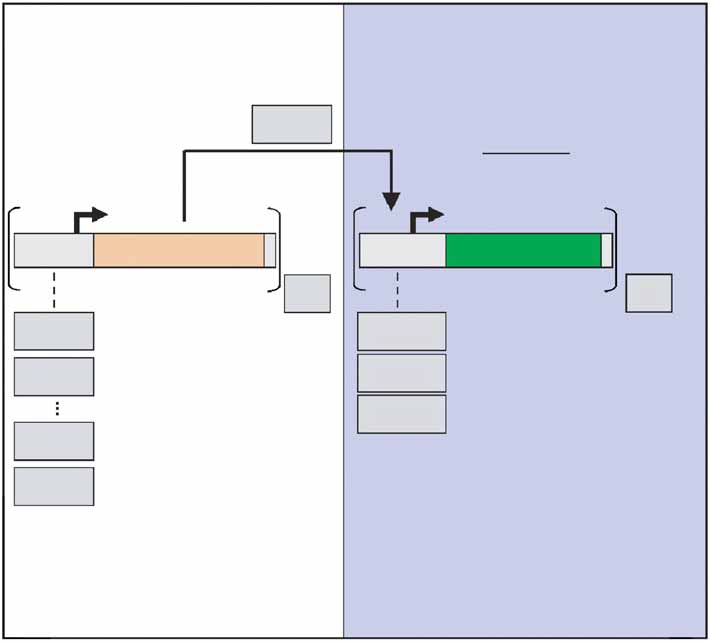
Figure 1 Synthetic gene circuits for noise amplification and transmission.
binding sites. In addition to the steeper response, the presence of
(a) The input and output modules are integrated into various chromosomal
multiple binding sites substantially lowered the induction threshold of
locations in a diploid cell. Gray boxes denote the experimental variables: the
the promoter. Approximately 5 and 45 times lower doxycycline
input and response promoters, the copy numbers N and M of the respective
concentrations are needed for P
modules and the external doxycycline concentration. (b) The output noise
TETO2 and PTETO7, respectively, than
equals the ratio of the standard deviation s
2 to the mean x2 of YFP
TETO1 to reach half-maximal expression. Notably, the basal
expression distribution. (c) Output noise Z2 of response promoters with a Hill
expression of PTETO1 was higher than that of the other constructs.
number of nH ¼ 1 (blue lines) or nH ¼ 2 (black lines) was calculated using
Increased basal expression of promoters with a single binding site has
equation S2 (Supplementary Note online), Zg ¼ 0.2 included. The dashed
been described for other transcription factors as well27.
and full lines stand for input noises of Z1 ¼ 0.4 and Z1 ¼ 0.5, respectively.
As predicted, Z2 was intensified by cooperative binding and
The effect of basal transcription (b ¼ 0.05) on Z2 is shown by a solid red
decreased as the output signal (YFP expression) increased (Fig. 2b
line (n ¼ 1, Z1 ¼ 0.4). The difference between the output noises elicitedby the two input promoters (arrows) is bigger when the response promoter
and Supplementary Fig. 1 online). The maximal value of the output
is cooperative, nH ¼ 2.
noise was approximately double that of the input noise as measured
VOLUME 37 [ NUMBER 9 [ SEPTEMBER 2005 NATURE GENETICS
-YFP (fluorescence)
Output signal x
-CFP (fluorescence)
Output signal x
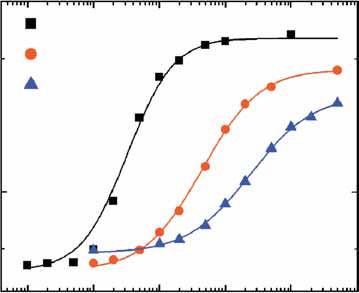
Figure 2 Amplification of noise by transcriptional cooperativity. (a,b) Output signal x2 and output noise Z2 of a single copy PTETO1-, PTETO2- or PTETO7-YFPconstructs driven by PSWI5-rtTA (ABY0529, ABY0530 and ABY0531). (a) The output signal was fitted to a Hill function with basal expression b, which is
enetics B0.05 times the maximal expression. (b) PCLN3-rtTA PTETO1-YFP (ABY0511a) with the relatively stronger PCLN3 was included to show that noise decreases at
high transcription rates. (c) Extrinsic and intrinsic (uncorrelated) noise in the noise amplifier system. Single-cell two-color fluorescence intensities are shownfor the strain ABY0528b (PSWI4-rtTA(S2) [PTETO7- CFP]1 / [PTETO7-YFP]1) using three different doxycycline concentrations (in mg ml–1). In the inset, the ratioof extrinsic to total noise is shown. (d) The output signal and noise of [PTETO7-YFP]1 was measured in the absence (empty symbols, ABY0535) and presenceof 14 additional [PTETO7-CFP]2 (filled symbols, ABY528a) rtTA binding sites. Symbols of the same shape denote induction with the same doxycycline
.com/natureg concentration (in mg ml–1).
directly by placing YFP under the control of PSWI5. In the case of
We tuned doxycycline concentrations to obtain equal levels of active
PTETO1, the Z2(x2) relation had a plateau. This phenomenon is
rtTA for a broad range of promoter strength (Fig. 3a). Among the
consistent with the above theoretical analysis of how the basal
examined promoters, PMYO2 had the lowest noise level (Fig. 3b,c).
http://www transcription rate affects noise.
PMYO2 drives constitutive expression of a cytoskeletal motor protein.
In contrast, PSWI6 had the highest noise level, roughly two times
oup Noise in the activity of cell cycle promoters
higher than that of PMYO2. Other cell cycle promoters, PSWI4, PSWI5,
Next, we examined how the presence of low-copy-number mRNAs
PCLN2 and PCLB2, had noise levels between those of PMYO2 and PSWI6.
affected the intensity of fluctuations in gene expression. Weak pro-
Because expression at PSWI6 is very low, it is expected to be affected
moters are inaccessible to direct noise measurement by fluorescent
substantially by intrinsic noise.
lishing Gr proteins. Even PSWI5-YFP produces a fluorescence that is only five
times greater than the cellular background fluorescence, although the
Fully correlated fluctuations at low expression level
SWI5 mRNA has B100 times higher copy number than the SWI6
To differentiate between intrinsic and extrinsic components of
mRNA. Therefore, we calculated the noise intensities Z1 by nonlinear
SWI6 noise, we examined how raising the input gene copy num-
regression of the experimentally determined amplified noise (Supple-
ber affected Z2. If noise in gene expression is purely intrinsic due
mentary Note and Supplementary Fig. 2 online). The output noise Z2
to the low copy number of transcribed mRNA, gene duplication
was determined predominantly by noise transmitted from the input
should decrease the output noise by a factor of
2 � 1:41, because
2005 Nature Pub
module Z1, because intrinsic noise in the response module was both
intrinsic noise is not correlated between different copies of a
low and approximately constant in the range of measured YFP
gene6. But when five copies of PSWI6-rtTA were integrated in tan-
fluorescence (Fig. 2c).
dem at the ade2 locus, the input noise Z1 was not reduced (Fig. 4a).
We used a triple integration of PTETO7-YFP to enhance the output
This suggests the absence of intrinsic, uncorrelated noise due to low
signal further, because the largest absolute differential amplification
copy number of expressed molecules. The five-copy construct had
of noise was observed at low intensities of the output signal. Theintroduction of 14 additional rtTA binding sites reduced the activepool of rtTA only slightly. This reduction in the number of activertTA did not increase the intensity of output noise, even when rtTA
was expressed from a weak promoter (Fig. 2d). Therefore, YFP
expression induced by rtTA reflected noise in the input module
faithfully; no additional noise was introduced by fluctuations in the
rtTA activity itself.
Output signal x2 (fluorescence)
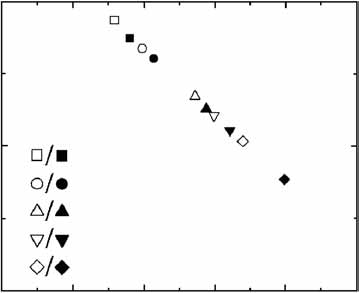
Figure 3 Input noise measurement using the noise amplifier system.
(a) Output signal of [PTETO7-YFP]3 with different input modules. (b) The
output noise Z2 for PCLB2-, PMYO2-, PSWI4- or PSWI6-rtTA and [PTETO7-YFP]3constructs. Solid lines denote fits by equation S2 (Supplementary Note
online). PCLB2-rtTA induces transcription even in the absence of doxycycline,
which accounts for the observed plateau in Z2(x2). For PSWI6-rtTA no data
were obtained at higher output mean values, because of the effect of
doxycycline on cell growth when its concentration is higher than 50 mg ml–1.
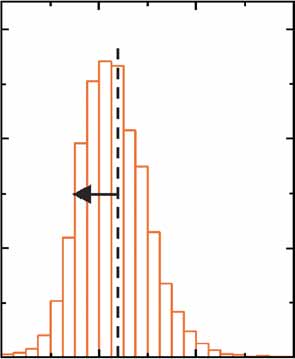
(c) Distribution of single-cell [PTETO7-YFP]3 fluorescence intensity, using the
MYO2-rtTA (ABY0520) or PBUD1-rtTA (ABY0548c) input modules at
Single-cell fluorescence
different doxycycline concentrations.
NATURE GENETICS VOLUME 37 [ NUMBER 9 [ SEPTEMBER 2005
ade2 /ade2
his3 /his3
Output signal x (fluorescence)
Output signal x (fluorescence)
Output signal x (fluorescence)
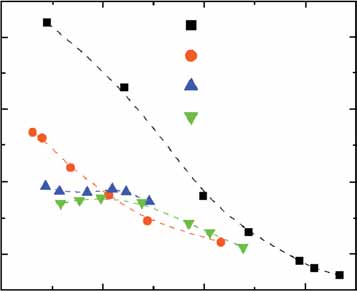
Figure 4 Dependence of fluctuations on the chromosomal position of P
SWI6. (a) The output noise Z2 for N ¼ 1, 2
and 5 copies of PSWI6-rtTA and [PTETO7-YFP]3 constructs (ABY0519, ABY0543 and ABY0545). (b) Output signal of
enetics [PTETO7-YFP]3 with various PSWI6-rtTA input modules. Symbols are as defined in the other panels. (c) Output noise
of a single copy of P
ade2 /ade2
SWI6-rtTA integrated at the ade2 (ABY0519) or his3 locus (ABY0547b). (d) Output noise
resulting from two copies of P
SWI6-rtTA integrated at the his3 locus in tandem (2xhis3, ABY0547a) or homologous
(his3/his3, ABY0561) arrangement. (e) Correlation between expression level and noise of PSWI6-rtTA integrations.
The active rtTA concentration is proportional to the product of rtTA and doxycycline concentrations. Therefore, we
his3 /his3
used the doxycycline concentration required to induce 25% of maximal YFP expression (b) as an indicator of the
rtTA expression level.
a slightly higher output noise than the single-copy construct, which
expected factor of noise reduction when a twofold increase in gene
may reflect the fact that longer chromosomal insertions with
expression stems from the increased frequency of gene activation,
http://www multiple tandem gene copies have slower association rates with the
which indicates that noise at SWI6 originates mostly in the rare
frequency of gene activation.
When a pair of PSWI6-rtTA constructs was integrated into homo-
logous chromosomal sites of a diploid cell, the input noise Z1 was
Chromosomal positioning affects correlation of fluctuations
reduced by a factor of 1.03 ± 0.01 relative to the single-copy
We observed a small reduction of noise when two copies of PSWI6-rtTA
integration (Fig. 4a). This reflects a strong but not full correlation
were integrated at homologous ade2 loci (Fig. 4a). To explore further
lishing Gr (r ¼ 0.88 ± 0.04) between two copies of a gene when integrated into
the effect of chromosomal positioning of genes on fluctuations, two
separate chromosomes. A correlation coefficient of r ¼ 1 indicates a
copies of PSWI6-rtTA were integrated either in tandem at a his3 locus
full correlation between individual events of promoter activation at
or one copy was integrated at each homologous his3 locus. The
different copies of a given gene. The full correlation of fluctuations in
resulting strains had very similar mean expression rates (Fig. 4b),
expression of tandem gene copies indicates that noise is not due to the
but the noise associated with the homologous pair was lower by a
low number of rtTA mRNA molecules but is generated in an event
factor of 1.09 ± 0.02 than that of the single-copy integration (Fig. 4d).
upstream of mRNA expression.
In contrast, the tandem integration did not decrease the noise. These
2005 Nature Pub
data indicate that the events of promoter activation are fully correlated
The effect of the frequency of promoter activation on noise
at genes integrated in tandem at a given chromosomal locus. When
The absence of intrinsic noise due to the low copy number of expressed
copies of a gene were positioned on separate homologous chromo-
molecules can be explained by a stochastic model of gene expression. In
somes, the correlation was reduced to r ¼ 0.68 ± 0.06, which is still
such a model, if a promoter is rarely activated and the rate of promoter
significant. We calculated the uncorrelated noise from the correlation
activation and inactivation is slower than the mRNA production rate
coefficient. Values of ZUNCOR were 0.14 ± 0.05 and 0.16 ± 0.05 for the
at an activated gene, then noise originates in the rare instances
homologous ade2 and his3 loci, respectively, which reflects the fact that
of transcription. The number of mRNAs transcribed at a single event
the noise originating in the uncorrelated fluctuations between homo-
of promoter activation has a negligible effect on noise in this case.
logous chromosomal loci were similar for both the ade2 and his3 loci.
Thus, if the relatively high noise of SWI6 is not due to the low copy
Plotting the calculated (input) noise of various PSWI6-rtTA integra-
number of the expressed mRNA, then it might originate in the low
tions as a function of rtTA expression level showed that noise was
frequency of random gene activation. This model predicts that increas-
independent of rtTA mRNA level (Fig. 4e). Noise was determined
ing the frequency of promoter activation leads to reduction of noise.
primarily by the chromosomal-position dependent frequency of geneactivation and to a lesser degree by fluctuations uncorrelated between
Chromosomal positioning changes expression level and noise
PSWI6 has no known transcriptional regulation, because it lacks aTATA box or binding site for any known transcriptional regulator28,29.
Propagation of fluctuations in single cells
The expression rate of several genes depends strongly on the chro-
These results shed light on the origins of fluctuations in concentration
mosomal or intranuclear positioning of the gene30. Therefore, we
of regulators. Next, we studied how regulator noise is propagated to
changed the chromosomal position of PSWI6-rtTA to examine whether
target genes. Although cooperative binding of transcription factors
this leads to a change in the rate of gene expression. When a single
increased the sensitivity in response, it also made the response noisier
copy of PSWI6-rtTA was integrated at the his3 locus, its expression rate
(Fig. 2b). To explore how fluctuations weaken the correlation between
doubled and was comparable to that of the two-copy integration at
the input and output signals, we measured the expression of an rtTA–
the ade2 locus (Fig. 4b). Noise at his3 was reduced substantially, by a
cyan fluorescent protein (CFP) fusion protein driven by the stronger
factor of 1.37 ± 0.04 relative to the ade2 locus (Fig. 4c,e), which is
PSWI5 promoter (Fig. 3a) and that of the PTETO7-YFP construct in
close to the maximal potential reduction (factor of 1.41). This is the
single cells (Fig. 5a). We plotted the output fluorescence (YFP) as a
VOLUME 37 [ NUMBER 9 [ SEPTEMBER 2005 NATURE GENETICS
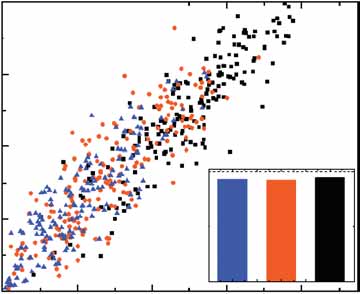
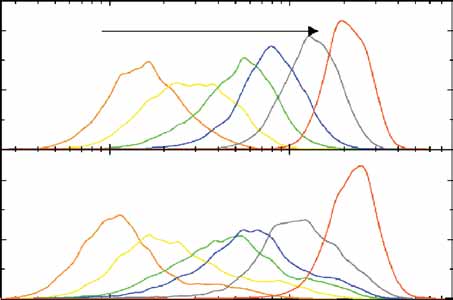
Figure 5 Correlation of fluctuations between the input and output modules.
(a) Two-color fluorescence imaging of rtTA-CFP (red) and P
TETO7-YFP (green)
in ABY0557 cells. (b) Correlation of input-output fluorescence intensitiesin single cells (ABY0557). The input fluorescence values (rtTA-CFP) weremultiplied by the ratio of the actual to the lowest doxycycline concentrationused (0.005 mg ml–1). The basal output signal (PTETO7-YFP) is 752,nH ¼ 2.15 when fitted to single-cell fluorescence data. Fitting the Hillfunction to the mean output signal when the doxycycline concentration isvaried yields nH ¼ 2.08 (data not shown). (c) Fluorescence of PSWI5-rtTA-YFP and PSWI5-rtTA-CFP constructs (ABY0559), integrated at homologousade2 loci, each having three copies. The total and uncorrelated noises were0.26 ± 0.03 and 0.15 ± 0.02, respectively. (d) The output signal inABY0549 is the fluorescence intensity of PTETO7-CFP. The input signal
enetics corresponds to PSWI5-YFP fluorescence, normalized by doxycycline
concentration as in b. nH ¼ 1.8 when fitted to single-cell fluorescence data.
(e) Normalized deviations for ABY0557 (black columns) and ABY0549 (red
columns). Doxycycline concentrations of 0.01 and 0.045 mg ml–1 were used
for respective strains to obtain similar input and output noise values for the
2dev4 ¼ 0.25, Z1 ¼ 0.262, Z2 ¼ 0.443 for ABY0557;
ox2dev4 ¼ 0.43, Z1 ¼ 0.292, Z2 ¼ 0.46 for ABY0549.
A-CFP fluorescence
(YFP fluorescence)
function of CFP fluorescence normalized by the applied doxycycline
Normalized input signal x1
rtTA-YFP fluorescence
concentration (Fig. 5b). Thus, the concentration of active rtTA can be
http://www correlated with the output signal in single cells, assuming a linear
relation between doxycycline and active rtTA concentration14. We
oup obtained the single-cell response obtained by fitting a Hill-type
function to the normalized input and response fluorescence data
obtained at three different doxycycline concentrations. The Hill
coefficient, obtained from single-cell measurements, was similar to
(CFP fluorescence)
lishing Gr that obtained from population-level measurements. Single-cell
measurements showed that even though the output signal was affected
Normalized input signal x
Normalized deviation
by large fluctuations, the input and output signals were significantlycorrelated in individual cells (Fig. 5b). Cells with lower rtTA expres-sion had lower output signals in the sigmoid region of promoter
were transmitted to target genes. Promoters that contain multiple
response and vice versa. To quantify the deviation from fully corre-
binding sites for Swi6p are the most likely to be affected by trans-
lated response, we calculated the average difference between single-cell
mitted noise, because chromatin-mediated cooperativity enhances the
2005 Nature Pub
output signals and the fitted response based on the Hill-type function,
propagation of fluctuations. The promoter of the gene BUD1, which
ox2dev4. x2dev equals the absolute value of (x2 – x2cal)/x2cal, where x2
regulates bud site selection, contains three closely located Swi4p-Swi6p
denotes the output signal in a single cell and x2cal denotes the expected
binding sites28. Noise associated with PBUD1 had a high intensity,
output value based on the fitted Hill-type function. We found that
comparable to that of PSWI6 (Figs. 3c and 6a). To link SWI6 and
ox2dev4 ¼ 0.25 for the above strain. In comparison, we found a
BUD1 fluctuations directly, we placed the expression of SWI6 under
value of 0.43 for a control strain, where the input signal was not
the control of PSWI5, which has roughly half the noise level of PSWI6
measured directly by an rtTA-fluorescent protein fusion but by PSWI5-
(Fig. 3b). This is expected to reduce the fluctuations in Swi6p
YFP integrated at homologous chromosomal location with respect to
concentration. Noise in PBUD1 activity was reduced substantially, by
PSWI5-rtTA (Fig. 5d). Stochastic expression from the PSWI5-YFP and
a factor of B2 (Fig. 6b). These observations indicate that noise at
PSWI5-YFP constructs was not fully correlated because they were
PBUD1 was predominantly transmitted from PSWI6.
positioned on physically distinct chromosomes, even though they
PCLN2 contains binding sites for Swi6p and for several additional
were driven by the same promoter (Fig. 5c). This accounts for the
transcriptional factors. The relatively lower noise level at PCLN2
larger deviation between the input and output signals. In summary, we
might be a result of regulation from multiple transcriptional
found a significant correlation between fluctuating concentrations of
factors (Fig. 6a).
the transcription regulator and its target gene. In principle, a strongcorrelation may be absent in some cases of eukaryotic gene activation.
A lower correlation is expected, for example, if independent stochastic
Fluctuations in transcription factor activity were amplified through
transitions between accessible and inaccessible states of a promoter
chromatin-mediated cooperativity (Fig. 2b), which could affect func-
influence the efficiency of transcription factor binding.
tioning of cellular regulatory networks because eukaryotic promotersusually contain multiple transcription-factor binding sites. Even
Transmission of SWI6 fluctuations to target genes
though the response signal was more broadly distributed over a cell
These results suggest that fluctuations in the concentration of reg-
population, our findings indicate that the input and response signals
ulatory proteins have a dominant role in determining noise strength in
of cooperative promoters were correlated in individual cells (Fig. 5b).
target genes, which is further amplified by the sensitivity in the
In principle, signal amplification through cooperative binding enables
promoter response. Next, we examined whether fluctuations in the
an efficient propagation of oscillations in individual cells, but the
concentration of Swi6p protein, inherited from the SWI6 promoter,
coamplified extrinsic noise could desynchronize cell-cycle oscillations
NATURE GENETICS VOLUME 37 [ NUMBER 9 [ SEPTEMBER 2005
Normalized noise 0.2
Normalized promoter strength
Output signal x (fluorescence)
Figure 6 Noise intensities of cell-cycle promoters. (a) Relation of noise topromoter strength. Promoter strength was determined using promoter-lacZ14
enetics or promoter-GFP constructs and was normalized by the promoter strength of
PSWI5 (compare with B1 copy per cell for SWI5 mRNA11). The input noisewas determined as in Figure 4e. For TETO2, noise was measured directly byinducing the PCLN3-rtTA:: [PTETO2-YFP]1 construct with high concentration ofdoxycycline (50 mg ml–1) to represent noise at saturated promoter activity.
.com/natureg Noise at PCLN2 is reduced in SWI4/swi4D heterozygote strains (from an
intensity of 0.294 ± 0.021 in wild-type to 0.265 ± 0.019), which indicatesthat part of the noise at PCLN2 is extrinsic due to regulation by Swi4p-
Figure 7 Model of noise generation. Gene activity can be strongly influenced
.nature Swi6p. (b) Output noise of PBUD1 is reduced when SWI6 is expressed under
by chromosomal position (blue and red circles stand for different positions;
the control of PSWI5 (blue circles, ABY0551a, Z1 ¼ 0.24) in comparison to
e.g., the ade2 and his3 loci). Noise is primarily determined by the
a wild-type strain (red squares, ABY0548c, Z1 ¼ 0.47).
association rate of the gene with the transcriptional machinery (purple foci).
The width of the arrows denotes the chromosomal position–dependent
rate of gene activation and deactivation. The two adjacent circles on a
in an initially synchronized cell population. A related phenomenon
chromosome represent tandem gene integration. Expression of genes at
oup was observed in the circadian clock of zebrafish cells. Population-level
homologous sites is not fully simultaneous. Noise in regulator expression,
measurements showed a fast decay of biorhythmic oscillations after
generated by the above mechanisms (e.g., SWI6), is transmitted to targetgenes (e.g., BUD1).
the cells were placed in a dark environment. Individual cells oscillatedregularly, however, but with distributed phases and fluctuating per-
lishing Gr iod31. Stochastic effects may also be buffered by the robust functioning
of the cell cycle32,33. Perturbations of the cell-cycle network can
tion rates with the foci (such as ade2), and others have faster rates
weaken noise buffering or further intensify noise and consequently
(such as his3). Less frequent stochastic association results in larger
derail the coordination of cell-cycle activities34. For example, impair-
fluctuations. This model also implies that fluctuations are fully
ing the positive feedback in cyclin-dependent kinase activation leads
correlated when genes are inserted in tandem arrangement, because
to stochastic, quantized cycle times in fission yeast35.
they associate with the foci simultaneously. The association of homo-
We started the search for intrinsic noise by correlating noise to
logous chromosomal regions with these foci occurs at equal frequency
2005 Nature Pub
promoter strength, expecting that as the promoter became weaker the
(hence the equal noise) but not completely simultaneously. As a result,
noise would become correspondingly stronger. This showed that genes
the correlation in fluctuations between the two loci is diminished.
with very low expression (SWI6 and, to some extent, SWI4) had high
In spatially extended systems, not all uncorrelated fluctuations
noise level, whereas BUD1 was affected by transmitted fluctuations
equal intrinsic noise. Even spatially homogeneous extrinsic fluctua-
(Fig. 6b). Notably, genes reported to have copy numbers of B1
tions can result in inhomogeneous accumulations of reaction compo-
mRNA (e.g., SWI5) per cell had only moderate noise, slightly above the
nents in some simple chemical diffusion reaction systems5. In
noise strength associated with saturated promoter activity (Fig. 6a).
principle, such inhomogeneously dispersed regulatory components
The full correlation of gene expression from multiple-copy PSWI6
can generate uncorrelated extrinsic fluctuations. In single cells, spa-
constructs in single cells is notable, given the very low transcript
tially inhomogeneous distribution of regulators can generate uncor-
abundance and the fact that PSWI6 has no known transcriptional
related fluctuations in expression of two identical genes situated in
regulators28. These observations show that the large cell-to-cell varia-
different positions in the nucleus. Therefore, the uncorrelated fluctua-
tion in PSWI6 activity is not intrinsic noise due to the low number of
tions of homologous chromosomal regions could be considered local
expressed mRNAs. Large fluctuations may instead originate in the
rare, random instances of promoter activation leading to mRNA
Our results link stochastic behavior of transcriptional networks to
production36. In principle, promoter activation may be initiated by
two primary sources: the random, rare instances of gene (promoter)
fluctuations in concentration of general transcription factors or by
activation determined primarily by chromosomal positioning of a
changes in the accessibility of the promoter in a given chromosomal
gene (Fig. 4e) and noise transmission through transcriptional regu-
region to transcription factors. For example, transcriptional activity
lators (Figs. 5 and 6b). In higher eukaryotes, positioning of genes
correlates with the spatial positioning of chromosomal regions in the
along the chromosomes and in the nucleus probably exerts a strong
yeast nucleus, and coregulated genes may be clustered in the
influence on stochastic gene activation for the following two reasons.
nucleus37–39. Additionally, active genes associate with spatially dis-
With increasing complexity of eukaryotic organisms, transcription
persed foci of RNA polymerase II in mammalian cells40.
of the genome is more and more repressed by multiple layers of
The above experimental observations are consistent with a model in
inhibitory mechanisms41,42. Therefore, events of gene activation by
which fluctuations are determined by the random association of
overcoming the repression may occur rarely and randomly for some
chromosomal regions with dispersed foci of the transcriptional
genes. Such events may cause the large cellular variation in gene
machinery (Fig. 7). Some chromosomal regions have slower associa-
expression when gene reporters are positioned close to repressed
VOLUME 37 [ NUMBER 9 [ SEPTEMBER 2005 NATURE GENETICS
regions of chromosomes43. In addition, individual genes and compo-
by the following equation:
nents of the transcriptional and post-transcriptional processing
machineries have specific intranuclear spatial arrangement44,45. There-
fore, expression of genes may be, to some extent, temporally uncorre-lated, owing to their distinct spatial regulation. The above phenomena
The rate of transcription at an activated promoter is n E 10 min–1 (ref. 49). In
may contribute to the stochastic behavior of gene expression observed
comparison, d and m have smaller values (0.05–0.5 min–1). Assuming that the
in higher eukaryotes46,47.
instances of gene activation are very rare events at low level of transcription,l o 0.01 min–1. In this case, n 4 d and m 4 l, which simplifies the aboveequation to
Construction of plasmids and strains. We inserted KpnI-promoter-BamHI,
BamHI-rtTA-EcoRI, BamHI-YFP-EcoRI or BamHI-CFP-EcoRI sequences into
l t1=2mRNA + t1=2gene
pRS402, pRS306 or pRS303 vectors unless otherwise specified14. The PBUD1,
enetics P
CLN3, PSWI5 and PMYO2 promoter sequences correspond to 600-, 1,200-, 892-
1/2mRNA and t1/2gene denote the half-lives of the mRNA and the
and 677-bp regions upstream of the start codon of the respective genes. We
activated gene, respectively. Assuming further that d 4 m, noise is given by a
binomial distribution of active gene states: Z2 ¼ m=l. In this region of
TETO1 and PTETO7 by replacing the tet operator region in PTETO2
upstream of the CYC1-TATA region14. rtTA corresponds to the rtTA(S2)
parameter values, Z2 is inversely proportional to the frequency of gene
variant. The rtTA-CFP and rtTA-YFP fusion proteins are constructed from
activation and is independent of mRNA production rate (e.g., as the copy
.com/natureg sequences encoding nuclear localization signal–tagged rtTA, rtTA-SV40NLS-
number of the integrated gene is varied).
BglII, fused to a C-terminally positioned BamHI-CFP-EcoRI. We integrated the
Mechanisms of transcription have been proposed that are capable of
producing very-low-copy-number mRNAs, in such a way that the timing of
SWI5-SWI6(1–500 + stop codon) cassette into the SWI6 locus by linearizing
the sequence with BseRI. All yeast strains (Supplementary Table 1 online) are
mRNA bursts are fully correlated even between multiple nonidentical promo-
cogenic with a diploid W303 strain (MATa/a ade2-1 leu2-3 ura3 trp1-1 his3-
ters50. A hypothetical transcription machinery composed of the RNA poly-
11,15 can1-100). We determined the number of integrations by Southern
merase linked to the DNA polymerase could produce RNA in a fully
blotting by digesting genomic DNA with restriction enzymes that cut the
deterministic way. A single mRNA molecule will be transcribed from each
http://www integrated construct at a single site.
gene during the passage of the above hypothetical machinery through thechromosome at a given cell division cycle.
oup Growth conditions and flow cytometric and microscopic analyses. We grew
To describe how the random instances of gene activation are correlated
yeast cells in minimal medium (synthetic dextrose supplemented with the
between two copies of a gene, we introduced the stochastic variables P1 (m1,
appropriate drop-out solution) at 30 1C. Exponentially growing cells were
Z1,1) and P2 (m2, Z1,2) with mean and noise given in parenthesis for each gene.
induced with doxycycline for 5 h unless otherwise indicated. Cell division time
If a pair of gene copies is integrated in tandem or at homologous sites (m1 ¼
lishing Gr is B90–100 min in this regime. We stopped expression by transferring the
m2 and Z1,1 ¼ Z1,2 ¼ Z1,S), then the resulting stochastic process P1+2(m1 + m2,
culture on ice. Cell density was A600 nm of 0.2–0.5. We evaluated fluorescence
Z1,D) depends on the correlation coefficient r between P1 and P2.
intensities using a narrow gating for forward and side scattering so that the
Then, the correlation between the transcriptional bursts at the two gene
most represented population was selected (central region of the scatter
copies is calculated from the ratio of noise of two gene copies to that of a single
distribution). We measured at least 10,000 cells. The fluorescence distributions
of promoter-GFP and -YFP constructs were identical.
For microscopic analysis, we obtained data on a Nikon TE2000 inverted
2005 Nature Pub microscope using Metamorph (Universal Imaging) data acquisition software.
For each doxycycline concentration, we measured 150–300 cells. Cells were
r reflects the proportion of correlated noise with respect to total noise:
outlined automatically in MATLAB using only the phase-contrast image.
Rare cases where the outline did not follow the cell wall were manually
discarded. We then mapped cell regions to the YFP and CFP fluorescent
images from which the total and average fluorescence were determined. We
Therefore, the uncorrelated noise was calculated as Z
obtained average cell fluorescence by dividing the total cell fluorescence by cell
area and subtracted a constant camera background from each frame. The
Note: Supplementary information is available on the Nature Genetics website.
autofluorescence of yeast cells was B7 times higher using a CFP filter cube thanusing a YFP filter cube.
ACKNOWLEDGMENTSWe thank J. Pedraza, W. Tansey and M. Thattai for discussions. A.B. is a Long
Data analysis. For presentation of output noise data, measurements are shown
Term Fellow of the Human Frontier Science Program. This work was supported
for mean fluorescence intensities higher than two times the cellular background
by a grant from the US National Institutes of Health and a US National Science
fluorescence, x2bg ¼ 3.6 ± 0.3, Z ¼ 0.36 ± 0.35. We obtained the Z2(x2) relation
Foundation CAREER grant.
for parameter fitting by subtracting the residual global noise at maximalinduction, Z
COMPETING INTERESTS STATEMENT
¼ 0.2, from the measured output noise: Z2ðx2Þ2 ¼
The authors declare that they have no competing financial interests.
� Z2. We obtained the input noise Z
1 by fitting equation S2
(Supplementary Note online) to the Z2(x2) relation, with n ¼ 1.45 and K ¼ 30(Supplementary Note online). We obtained normalized values of Z
Received 11 February; accepted 21 June 2005
Published online at http://www.nature.com/naturegenetics/
dividing Z1 by 2.0. This normalization factor corresponds to the ratio of inputnoise extrapolated from measurement of the amplification method (Z1 ¼ 0.55;Fig. 3b) to the noise measured directly, Z ¼ 0.27, using the PSWI5-YFP
1. Rao, C.V., Wolf, D.M. & Arkin, A.P. Control, exploitation and tolerance of intracellular
construct, which had a mean fluorescence of 29.
noise. Nature 420, 231–237 (2002).
2. Becskei, A., Seraphin, B. & Serrano, L. Positive feedback in eukaryotic gene networks:
Correlation between fluctuations in gene activation. We modeled the
cell differentiation by graded to binary response conversion. EMBO J. 20, 2528–2535
production of mRNA by probabilistic transitions between active (A) and
inactive (I) states of a gene48. The rates of promoter activation (I-A) and
3. Thattai, M. & van Oudenaarden, A. Stochastic gene expression in fluctuating environ-
ments. Genetics 167, 523–530 (2004).
inactivation (A-I) are denoted by l and m, respectively. The mRNA produc-
4. Erdi, P. & Toth, J. Mathematical Models of Chemical Reactions: Theory and Appli-
tion at an active promoter (A-A + mRNA) proceeds at a rate of n, and mRNA
cations of Deterministic and Stochastic Models (Manchester University Press,
degradation (mRNA-+) at a rate of d. In this case, the mRNA noise is given
Manchester, UK, 1989).
NATURE GENETICS VOLUME 37 [ NUMBER 9 [ SEPTEMBER 2005
5. Horsthemke, W., Doering, C.R., Ray, T.S. & Burschka, M.A. Fluctuations and correla-
28. Harbison, C.T. et al. Transcriptional regulatory code of a eukaryotic genome. Nature
tions in a diffusion-reaction system - unified description of internal fluctuations and
431, 99–104 (2004).
external noise. Phys. Rev. A. 45, 5492–5503 (1992).
29. Basehoar, A.D., Zanton, S.J. & Pugh, B.F. Identification and distinct regulation of yeast
6. Elowitz, M.B., Levine, A.J., Siggia, E.D. & Swain, P.S. Stochastic gene expression in a
TATA box-containing genes. Cell 116, 699–709 (2004).
single cell. Science 297, 1183–1186 (2002).
30. Maillet, L. et al. Evidence for silencing compartments within the yeast nucleus: a role
7. Paulsson, J. Summing up the noise in gene networks. Nature 427, 415–418 (2004).
for telomere proximity and Sir protein concentration in silencer-mediated repression.
8. Pedraza, J.M. & van Oudenaarden, A. Noise propagation in gene networks. Science
Genes Dev. 10, 1796–1811 (1996).
307, 1965–1969 (2005).
31. Carr, A.J. & Whitmore, D. Imaging of single light-responsive clock cells reveals
9. Rosenfeld, N., Young, J.W., Alon, U., Swain, P.S. & Elowitz, M.B. Gene regulation at the
fluctuating free-running periods. Nat. Cell Biol. 7, 319–321 (2005).
single-cell level. Science 307, 1962–1965 (2005).
32. Lengronne, A. & Schwob, E. The yeast CDK inhibitor Sic1 prevents genomic instability
10. Ghaemmaghami, S. et al. Global analysis of protein expression in yeast. Nature 425,
by promoting replication origin licensing in late G(1). Mol. Cell 9, 1067–1078 (2002).
737–741 (2003).
33. Thornton, B.R., Chen, K.C., Cross, F.R., Tyson, J.J. & Toczyski, D.P. Cycling without the
11. Holland, M.J. Transcript abundance in yeast varies over six orders of magnitude.
cyclosome: modeling a yeast strain lacking the APC. Cell Cycle 3, 629–633 (2004).
J. Biol. Chem. 277, 14363–14366 (2002).
34. Magee, J.A., Abdulkadir, S.A. & Milbrandt, J. Haploinsufficiency at the Nkx3.1 locus.
12. Velculescu, V.E. et al. Characterization of the yeast transcriptome. Cell 88, 243–251
A paradigm for stochastic, dosage-sensitive gene regulation during tumor initiation.
Cancer Cell 3, 273–283 (2003).
enetics 13. Swain, P.S. Efficient attenuation of stochasticity in gene expression through post-
35. Sveiczer, A., Csikasz-Nagy, A., Gyorffy, B., Tyson, J.J. & Novak, B. Modeling the fission
transcriptional control. J. Mol. Biol. 344, 965–976 (2004).
yeast cell cycle: quantized cycle times in wee1–cdc25Delta mutant cells. Proc. Natl.
14. Becskei, A., Boselli, M.G. & van Oudenaarden, A. Amplitude control of cell-cycle waves
Acad. Sci. USA 97, 7865–7870 (2000).
by nuclear import. Nat. Cell Biol. 6, 451–457 (2004).
36. Pirone, J.R. & Elston, T.C. Fluctuations in transcription factor binding can explain the
15. Hasty, J., Dolnik, M., Rottschafer, V. & Collins, J.J. Synthetic gene network for
graded and binary responses observed in inducible gene expression. J. Theor. Biol.
entraining and amplifying cellular oscillations. Phys. Rev. Lett. 88, 148101 (2002).
226, 111–121 (2004).
.com/natureg 16. Isaacs, F.J., Hasty, J., Cantor, C.R. & Collins, J.J. Prediction and measurement of an
37. Menon, B.B. et al. Reverse recruitment: The Nup84 nuclear pore subcomplex mediates
autoregulatory genetic module. Proc. Natl. Acad. Sci. USA 100, 7714–7719 (2003).
Rap1/Gcr1/Gcr2 transcriptional activation. Proc. Natl. Acad. Sci. USA 102, 5749–
17. Basu, S., Mehreja, R., Thiberge, S., Chen, M.T. & Weiss, R. Spatiotemporal control of
5754 (2005).
gene expression with pulse-generating networks. Proc. Natl. Acad. Sci. USA 101,
38. Casolari, J.M. et al. Genome-wide localization of the nuclear transport machinery
6355–6360 (2004).
couples transcriptional status and nuclear organization. Cell 117, 427–439 (2004).
18. Xiong, W. & Ferrell, J.E., Jr. A positive-feedback-based bistable ‘memory module' that
39. Hurst, L.D., Pal, C. & Lercher, M.J. The evolutionary dynamics of eukaryotic gene order.
governs a cell fate decision. Nature 426, 460–465 (2003).
Nat. Rev. Genet. 5, 299–310 (2004).
19. Pramila, T., Miles, S., GuhaThakurta, D., Jemiolo, D. & Breeden, L.L. Conserved
40. Osborne, C.S. et al. Active genes dynamically colocalize to shared sites of ongoing
homeodomain proteins interact with MADS box protein Mcm1 to restrict ECB-
transcription. Nat. Genet. 36, 1065–1071 (2004).
dependent transcription to the M/G1 phase of the cell cycle. Genes Dev. 16, 3034–
41. Struhl, K. Fundamentally different logic of gene regulation in eukaryotes and prokary-
3045 (2002).
otes. Cell 98, 1–4 (1999).
oup 20. Acar, M., Becskei, A. & van Oudenaarden, A. Enhancement of cellular memory by
42. Martin, D.I. Transcriptional enhancers–on/off gene regulation as an adaptation to
reducing stochastic transitions. Nature 435, 228–232 (2005).
silencing in higher eukaryotic nuclei. Trends Genet. 17, 444–448 (2001).
21. Shibata, T. & Fujimoto, K. Noisy signal amplification in ultrasensitive signal transduc-
43. Ahmad, K. & Henikoff, S. Modulation of a transcription factor counteracts heterochro-
tion. Proc. Natl. Acad. Sci. USA 102, 331–336 (2005).
matic gene silencing in Drosophila. Cell 104, 839–847 (2001).
22. Hooshangi, S., Thiberge, S. & Weiss, R. Ultrasensitivity and noise propagation in a
44. Misteli, T. Concepts in nuclear architecture. Bioessays 27, 477–487 (2005).
synthetic transcriptional cascade. Proc. Natl. Acad. Sci. USA 102, 3581–3586
45. Roix, J.J., McQueen, P.G., Munson, P.J., Parada, L.A. & Misteli, T. Spatial proximity
lishing Gr
of translocation-prone gene loci in human lymphomas. Nat. Genet. 34, 287–291
23. Blake, W.J., Kaern, M., Cantor, C.R. & Collins, J.J. Noise in eukaryotic gene expression.
Nature 422, 633–637 (2003).
46. Dernburg, A.F. et al. Perturbation of nuclear architecture by long-distance chromosome
24. Raser, J.M. & O'Shea, E.K. Control of stochasticity in eukaryotic gene expression.
interactions. Cell 85, 745–759 (1996).
Science 304, 1811–1814 (2004).
47. Nutt, S.L. et al. Independent regulation of the two Pax5 alleles during B-cell
25. Li, G. & Widom, J. Nucleosomes facilitate their own invasion. Nat. Struct. Mol. Biol.
development. Nat. Genet. 21, 390–395 (1999).
11, 763–769 (2004).
48. Peccoud, J. & Ycard, B. Markovian modelling of gene product synthesis. Theor. Popul.
26. Vashee, S., Melcher, K., Ding, W.V., Johnston, S.A. & Kodadek, T. Evidence for two
Biol. 48, 222–234 (1995).
2005 Nature Pub
modes of cooperative DNA binding in vivo that do not involve direct protein-protein
49. Iyer, V. & Struhl, K. Absolute mRNA levels and transcriptional initiation rates in
interactions. Curr. Biol. 8, 452–458 (1998).
Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 93, 5208–5212 (1996).
27. Melcher, K. & Xu, H.E. Gal80-Gal80 interaction on adjacent Gal4p binding sites is 50. Guptasarma, P. Does replication-induced transcription regulate synthesis of the myriad
required for complete GAL gene repression. EMBO J. 20, 841–851 (2001).
low copy number proteins of Escherichia coli?. Bioessays 17, 987–997 (1995).
VOLUME 37 [ NUMBER 9 [ SEPTEMBER 2005 NATURE GENETICS
Source: http://qobweb.igc.gulbenkian.pt/courses/coimbrasysbio2009/NatureGenetics-2005-37-937.pdf
GROUPE SRIB 2013 RAPPORT ANNUEL AMBITIOUS BRUSSELS Numeca International Radionomy & MusicMatic RAPPORT ANNUEL 13 20 05Le mot du Président 06 STRUCTURE Le Conseil d'administration La lettre du Conseil La direction et les collaborateurs du groupe finance.brussels : un groupe dédicacé aux entreprises et à la Région de Bruxelles-Capitale Les chiffres-clés du groupe
El artículo 14 de la nueva Ley Peruana de Arbitraje: Reflexiones sobre el contrato de arbitraje – realidad Eduardo Silva Romero* 1. Quienes elaboran las normas jurídicas deben, en términos generales, mantener un equilibrio (bastante frágil) entre los valores de la flexibilidad (y, por ende, adaptabilidad) del Derecho a situaciones futuras y la previsibilidad del mismo;


